Seaweeds can often be found along the borders between land and sea; better known as the coast. Seaweed can be found in these coastal locations as either either have permanent inhabitants, such as the intertidal seaweeds, or beach debris where pieces of seaweeds are washed ashore by waves and tides.
Green (Ulva spp) and brown (Fucus spp) seaweed at low tide. Saltdean. East Sussex. UK.
As borders between land and sea, coastal locations can often be important transitional zones for animals to supplement their diet. Polar bears, wolves and brown bears are well known to appreciate the bounty of a stranded whale carcass washed upon the shore (Lewis & Lafferty, 2014; Laidre et al., 2018).
It is not just carnivores that benefit from the sea’s bounty though. In the isolated Svalbard archipelago, located 800km north of mainland Norway in the middle of the Arctic Ocean, well north of the Arctic Circle, coastal seaweed is becoming an important resource in the face of climate change.
Longyearbyen. Svalbard.
Warming temperatures in the archipelago are creating more frequent rain on snow conditions, whereby thick layers of ice cover the land vegetation, including mosses and lichens. These impenetrable ice-locked pastures make foraging for food difficult for Svalbard’s herbivorous animals. Including the Arctic’s most famous grazer, the reindeer.
Reindeer. Svalbard.
Svalbard’s reindeer (Rangifer tarandus platyrhynchu) is a peculiar type, thanks to its unique habitat. They are shorter and stockier than mainland European and north American reindeer, being well suited to the cold of the far north. Unlike their mainland relatives Svalbard reindeer do not show migratory behaviour, instead they could be considered rather lazy, being characterized by a stationary, energy‐saving lifestyle. This lifestyle is only possible in Svalbard by virtue of the unusual lack of predation. The Svalbard reindeer population is instead controlled by climate and density dependent processes, with starvation being the most common cause of death.
Reindeer. Svalbard.
With the availability of food being a very important factor effecting survival, the increasing occurrence of ice-locked pastures could pose quite fatal for Svalbard reindeer.
But some have come up with a clever coping mechanism. On the northwestern coast of Spitsbergen, the largest island in the Svalbard archipelago, 13% of the total reindeer population turned to the coast for food (Hansen and Aanes, 2012). They were found to developed a taste for eating kelp and other seaweeds that had been washed ashore.
Svalbard reindeer eating seaweed. Photo: Brage B. Hansen/NTNU.
However the reindeer cannot sustain themselves entirely on seaweed, being observed to frequently move between beaches and more accessible pastures. The seaweed scraps were being used as an exotic supplement to the reindeer’s normal plant‐based diet.
By adjusting their behavioural the reindeer utilise resources of sea origin to help survival during harsh winters. With the prediction of far more frequent harsh winters due to climate change, seaweed could act as a vital buffer in the face of environmental variation.
And Svalbard reindeer are not unique, many other herbivores eat kelp or seaweed too. From the sheep (Ovis aries) on Orkney [Scotland] (Hall, 1975), red deer (Cervus elaphus) on the Isle of Rum [Scotland] (Conradt, 2000), to black‐tailed deer (Odocoileus hemionus) of channel island [Alaska] (Parker et al., 1999).
Sheep feeding on seaweed along the shoreline. North Ronaldsay. Orkney. Scotland.
So seaweed is not just important in the sea world, it could be helping many land species too, especially in the face of climate change.
This post was based on the research of Hansen et al., (2019). If you enjoyed this post you can find the open access article here:
Conradt, L. 2000. Use of a seaweed habitat by red deer (Cervus elaphus L.). Journal of Zoology 250:541–549.
Hall, S. J. 1975. Some recent observations on Orkney sheep. Mammal Review 5:59–64.
Hansen, B. B., and R. Aanes. 2012. Kelp and seaweed feeding by High‐Arctic wild reindeer under extreme winter conditions. Polar Research 31:17258.
Laidre, K.L., Stirling, I., Estes, J.A., Kochnev, A. and Roberts, J., 2018. Historical and potential future importance of large whales as food for polar bears. Frontiers in Ecology and the Environment 16(9), pp.515-524.
Lewis, T.M. and Lafferty, D.J., 2014. Brown bears and wolves scavenge humpback whale carcass in Alaska. Ursus, International Association for Bear Research and Management 25(1), pp.8-13.
Parker, K. L., M. P. Gillingham, T. A. Hanley, and C. T. Robbins. 1999. Energy and protein balance of free‐ranging black‐tailed deer in a natural forest environment. Wildlife Monographs 143:3–48.
Fucus is the scientific name for the group of seaweeds that bladderwrack belongs. They are large, brown seaweeds that can live for many years. Typically residing in the intertidal zones of rocky seashores this group can be found throughout much of the northern hemisphere. The members of Fucus are commonly called to as wracks.
An underwater Fucus) forest at Brofjorden. Sandvik. Sweden.
Their widespread distribution and the large number of species found residing in a tight zone has led to them being sought after seaweeds to be used in the study of many scientific concepts including zonation and competition.
Within the Baltic Sea four species are found, three native and one invasive. By native we mean that their North Sea ancestors entered the Baltic Sea naturally around 8000 years ago and they have been resident ever since (Ardehed et al., 2016). Whereas the invader was introduced to the enclosed sea ~100 years ago, possibly by human activity (Wikström, 2004). This invader is an alien to the Baltic Sea ecosystem and that could potential cause a harmful impact on the otherwise native community.
Tiny wrack (Fucus spp.) surrounded by barnacles.
The Baltic Sea is not the typical habitat of Fucus, and that’s a big reason why we find fewer species then you might expect on a normal coastline. Wracks are marine species with those inhabiting areas outside of the Baltic Sea normally living in true marine conditions. However all wracks in the Baltic Sea have some tolerance to the lower salinity conditions present in this semi-enclosed sea. This tolerance has a huge impact on where we can find each species. Each species can tolerate a different salinity range, with those that can tolerate lower ranges being found deeper into the inner Baltic Sea than those who can’t.
But who are these four Fucus of the Baltic Sea?
The most famous, and most widely spread of them, as you might guess, is bladderwrack. Bladderwrack can be found along most of the European coastline and throughout the Baltic Sea until the far northern parts of the Gulf of Bothnian and the eastern parts of the Gulf of Finland. Bladderwrack is able to tolerate a range of salinity from true marine conditions down to truly brackish conditions of the Baltic Sea. This is why we find it so extensively.
Bladderwrack (Fucus vesiculosus).
The second species, has a far narrower tolerance range, and as a species of marine origin it cannot be found when salinity gets too low. The typical form has a saw-like appearance, hence it’s name: serrated wrack. You’ll find serrated wrack all along the west coast of Sweden and along the southern east coast, but not quite as far north as Stockholm. Serrated wrack can also be found on the Swedish islands of Öland and Gotland.
Serrated wrack (Fucus serratus ).
Up until quite recently, these two species were thought to be the only two native wracks. The extensive range of bladderwrack and fairly limited one of serrated wrack is the reason why for many years bladderwrack was viewed as the one and only species within the majority of the Baltic Sea. Accordingly, if you were in Finland or Estonia and you found some wrack, it was certainly bladderwrack.
However this might not be the case… In 2005, it all changed. A group of researchers from Sweden used the same techniques that I use within my own research to throw all previous assumptions up in the air. Enter Narrow wrack…
Narrow wrack (Fucus radicans). Image from Tangbloggen.
For many years prior to this, researchers were commenting on a strange dwarf form of bladderwrack found in the Baltic Sea. This was assumed to be a morphotype (a group of distinctly different looking individuals within the same species) of bladderwrack. But by looking at the DNA of this dwarf the researchers found that it was not only morphologically distinct and but also genetically distinct (Bergstöm et al., 2005). This morphotype was afforded species level status. Thus now if you are in the northern parts of the Baltic Sea, in either Sweden or Finland, or even on the Estonian island of Saaremaa and find some wrack you could be looking at either bladder or narrow wrack.
Narrow wrack diverged from the same North Sea ancestors as those of Baltic Bladderwrack shortly after they entered the Baltic Sea (Ardehed et al., 2016). As such, narrow wrack is considered an endemic species to the Baltic. This means that it is both a native species to the Baltic Sea, but also it won’t be found anywhere else either.
An illustration of where you can find each Fucus within the Baltic Sea. The distribution of narrow wrack, bladderwrack, and serrated wrack are denoted by orange, green, and yellow respectively. Image from Stockholm University DEEP.
So these are the three natives of the Baltic Sea: Bladderwrack, serrated wrack and narrow wrack. But who is the invader?
Well this invader doesn’t have a readily available English common name but in Finnish it’s called häilyhauru. Perhaps not the easiest to pronounce so instead I will use it’s scientific name: Fucus evanescens. In fact this wrack is actually of dubious identity, potentially being either a species in its own right or a subspecies a different rockweed. Either way this seaweed invaded many European coasts including the Baltic Sea from its origins in the Arctic. Perhaps surprisingly, the invasion of Öresund at the entrance to the Baltic Sea in 1966-72 was very well documented 1966-72 (von Wachenfeldt 1975). We know now that this wrack can be found throughout the Danish straits, Swedish sound, and the Baltic German coastline.
Häilyhauru (Fucus evanescens ).
So now we have it, the four Fucus of the Baltic Sea. They are important components of the Baltic Sea ecosystem, providing home and food for many other plants, animals, and seaweeds. Without these wrack the Baltic Sea would be a very different place, and more importantly I would be out of a research topic!
Sources
Ardehed, A., Johansson, D., Sundqvist, L., Schagerström, E., Zagrodzka, Z., Kovaltchouk, N.A., Bergström, L., Kautsky, L., Rafajlovic, M., Pereyra, R.T. and Johannesson, K., 2016. Divergence within and among seaweed siblings (Fucus vesiculosus and F. radicans) in the Baltic Sea. PLoS One, 11(8), p.e0161266.
Bergström, L., Tatarenkov, A., Johannesson, K., Jönsson, R.B. and Kautsky, L., 2005. Genetic and morphological identification of Fucus radicans sp. Nov.(Fucales, Phaeophyceae) in the brackish Baltic Sea 1. Journal of Phycology, 41(5), pp.1025-1038.
von Wachenfeldt T (1975) Marine benthic algae and the environment in the Öresund. Systematic Botany, Lund University:328.
Wikström, S.A., 2004. Marine seaweed invasions: the ecology of introduced Fucus evanescens (Doctoral dissertation, Botaniska institutionen).
When I first came to Finland and started working with Baltic Sea bladderwrack I knew nothing about this unique sea. But if you ignore the fact that the Baltic Sea is so very different than the typical seas then you risks your ideas on the biology of any Baltic Sea plant animal or algae being wrong. So this month lets look at why the Baltic Sea is so unique.
Furuskär, a Baltic Sea island in the Tvärminne archipelago.
The most defining feature of the Baltic Sea is the low salinity. In simple terms the salinity is the saltiness of the water. In typical seawater you would expect a salinity of 30-35‰ whereas in the Baltic Proper you will not experience salinity any higher than around half this value. Despite being called a sea, it does not possess the characteristic salty seawater you’d expect from that name. But though the Baltic Sea is not like typical seawater, it is also not like typical ponds, lakes or river either. In fact it’s waters lies somewhere in between the two, being not quite sea or freshwater either. The easiest way to describe it is brackish, a sort of halfway water, with the average salinity being just ⅕ of typical seawater (Leppäranta & Myrberg, 2009).
But does this really make such a difference? Well it certainly does!
Three-spined stickleback (Gasterosteus aculeatus). A common fish found within the Baltic Sea with a widespread distribution throughout the Northern Hemisphere in both fresh and salt water.
Plants, animals and algae have preferred environmental conditions that they can tolerate. This can be considered a tolerance range. Most of us like the summer heat of 30°C and don’t mind a winter of -1°C, but if we were then forced to live in 70°C or -60°C we probably wouldn’t last long. So we can tolerate the moderate temperatures, but the extremes are too much to survive. Well this is the same for salinity. For example, marine fish do well within seawater and lake fish likewise in freshwater, but if you took a lake fish to the sea or a sea fish to the lake they would not be able to endure the different conditions. Each is adapted to tolerate the habitat that they live in.
Some of you might be wondering about some of the rather high profile fish that move between the sea and rivers (ahem..salmon..). Well it is true that some fish can move between salinity conditions, however for the most part they must slowly acclimatise to the new conditions undergoing changes to their bodies to allow them to tolerate the new environmental conditions. Acclimatising to extremes is possible, but not very common. Generally, most plants, animals, and algae have a range they can tolerate and if subjected to conditions outside of this then the results end badly.
Atlantic salmon (Salmo salar) heading up the Tyne River. UK.
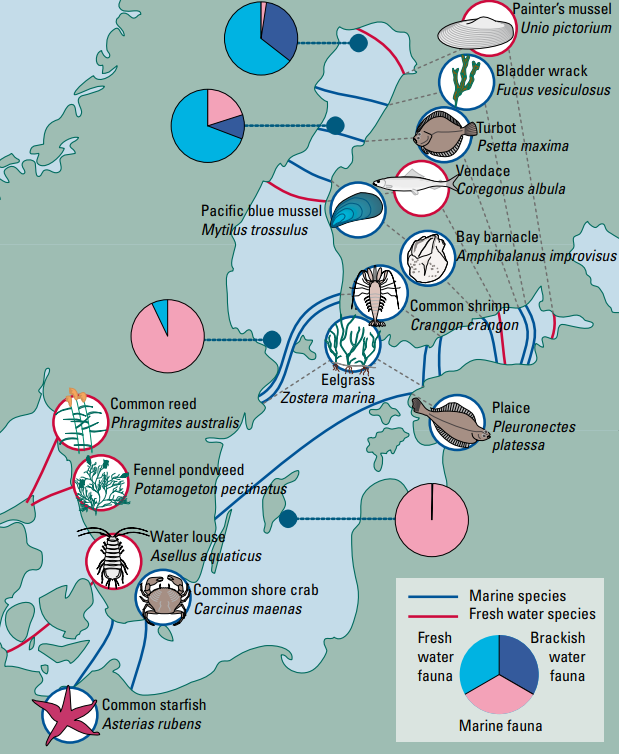
So back to the Baltic Sea. As a brackish sea, marine life are out of their comfort zone, but so too are the freshwater species. This means that many marine species that you would find around the coasts of Europe cannot live in the Baltic Sea just as freshwater species in continental Europe cannot either. This results in species paucity: only a few inhabitant species. In fact the adjacent North Sea has ~10X more species than the Baltic Sea (Elmgren and Hill, 1997). Characteristic marine organisms such as starfish and sea urchins are missing from most of this sea, though other species that you wouldn’t expect from a sea which are pretty abundant such as the common reed.
Example distribution of plants, animals, and seaweeds in the Baltic sea. Notice that marine animals become less common in northern areas whereas freshwater animals are absent from the southern areas. (Furman et al., 2014).
In Fucus terms; the group of seaweeds that bladderwrack belongs to; you could expect to find anywhere up to 3 or 4 species on a typical British rocky shore, whereas in the Baltic Sea you would be lucky to find more than one. In fact, if you searched throughout the whole Baltic Sea you’d only find three native Fucus species: bladderwrack, narrow wrack and serrated wrack.
Bladderwrack (Fucus vesiculosus). Torslanda. Västra Götaland County. Sweden.
Though the salinity within the Baltic is always brackish, not every location is the same. In fact the salinity ranges from (20-)10‰ (depending on where you decide the Baltic Sea starts) down to less than 1‰ occurs (Waern, 1952). This range is actually quite predictable, forming rather well defined gradients.
But why do we see this gradient?
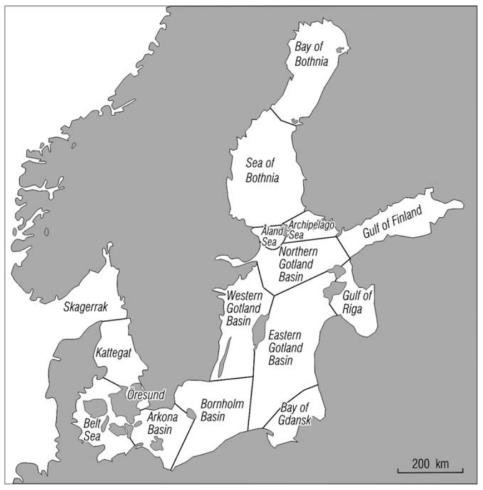
Salt water can only flow into the Baltic Sea from one relatively narrow location in the south over the Danish belts and the Swedish sound. These channels are shallow, narrow sills that restrict the flow of saltwater into the Baltic Sea. Because seawater is denser than fresh and brackish water it does not readily transfer over these shallow sills, with only a few major inflows happening per year. This means that the input of high salinity seawater into the Baltic Sea is rare. To add to this, the Baltic Sea is surrounded by land. This area of land surrounding the Baltic Sea that drains water into the sea is four times larger than the sea itself (Zillén et al., 2008). From this large area of land, freshwater can runoff into the Baltic Sea and consequently reduces the salinity. The largest single input of freshwater occurs in the Gulf of Finland by the river Neva, equating to 15% of the total Baltic river inflow (Alenius et al., 1998; Kuosa and Myrberg, 2009). Other large river inputs include the Vistula, the Daugava, the Nemunas, the Kemijoki, the Oder and the Göta Älv (Furman et al., 2014). Because the large rivers occur in the north or east and the only saltwater input comes from the south a well-defined south-north salinity gradient is produced.
Map of the Baltic Sea and its sub-basins. The three Danish straits and Swedish Sound (Little Belt, Öresund, and Storebælt) are located in the south west in the Belt Sea and Öresund. (Leppäranta and Myrberg, 2009).
So this all means is that the Baltic Sea is a semi-enclosed brackish system, where high salinity water is only provided in the south and freshwater input from the land dilutes the sea into the brackish conditions we see. But how long does water stay in the Baltic? Well it takes approximately 40 years for Baltic Sea water to be renewed (Leppäranta & Myrberg, 2009).
The Baltic Sea is not the only environment to be considered brackish, saltmarshes and estuaries can also frequently be considered brackish. These environments share similar characteristics of salinity, however the Baltic Sea has many other features to add to its uniqueness.
A typical saltmarsh.
The Baltic Sea has a rather characteristic, elongated shape, ranging approximately 1,300km from north to south and 1,000km from west to east covering an area of 412,560km2 (Gerlach, 1994; Seifert & Kayser 1995). The sea actually lies across climatic zones, covering both maritime temperate zones and continental sub-arctic climate zones (Leppäranta & Myrberg, 2009). That means that different locations throughout the Baltic Sea will experience differing climatic conditions. Since the Baltic Sea reaches into the sub-arctic zone combined with the brackish water, ice is a common occurrence in winter. During the winter months some 40% of the Baltic Sea is covered in ice, with the greatest amount being seen between January and March (Finnish Meteorological Institute, 2017a).
A view of the Baltic Sea from space. Sweden appears on the left and Finland in the upper right. The sea ice envelops the coastal islands with sprinkles of sea ice away from the coast. Credit: NASA image courtesy the MODIS Rapid Response Team at NASA GSFC.
The deepest part of the Baltic Sea is at Landsort Deep in the Baltic Sea Proper, reaching a maximum depth of 459m (Furman et al., 2014). Maybe this sounds pretty deep, but when you compare it to the Atlantic and Pacific Oceans with a maximum depth of 8710m at Milwaukee Deep or ~10900m at Challenger Deep respectively then it seems pretty shallow (Stewart & Jamieson, 2019). In fact the mean depth of the Baltic Sea is a measly 54m, that’s just over the length of an Olympic-size swimming pool (Furman et al., 2014). Moreover much of the Baltic Sea lies well above this depth, with the Gulf of Finland averaging 37m (Kuosa and Myrberg, 2009) and the Gulf of Riga just 27m (Szaniawska, 2018).
Saaremaa. Estonia. Gulf of Riga.
Despite its elongated shape, the shallow depth means that the total volume of the Baltic Sea is merely 21,631km3 (Seifert & Kayser, 1995); that equates to <0.1% of the total ocean volume on earth (Eakins & Sharman, 2010). So the Baltic Sea is just a drop in the ocean when compared to the Atlantic (23.3%) and the Pacific (49.4%) oceans.
Though being inconsequential in the grand scheme of the global oceans, the Baltic Sea has a huge effect on and is also hugely affected by human activities. Nine countries surround the semi-enclosed sea: Denmark, Germany, Poland, Lithuania, Latvia, Estonia, Russia, Finland and Sweden. The sea has been integral in the development of these countries and has supported many a livelihood.
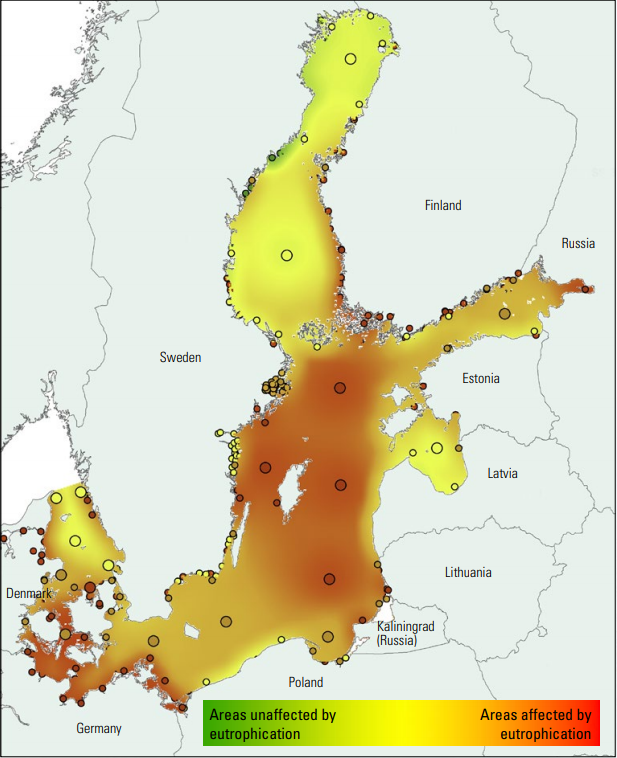
Eutrophication is a negative impact on the Baltic Sea caused by human activity whereby excess nutrients are leaked into the sea. This can have many bad effects on the Baltic Sea, and has caused mass die-offs of bladderwrack in the past. Notice that most of the Baltic Sea is affected by eutrophication, with much of it being quite severe. (HELCOM 2010a).
To add to the peculiarity of the Baltic Sea, tides are absent. In the Baltic Sea you will not experience the daily fluctuation of seawater traveling up and down the shore. For plants, animals, and algae living on the shore this means that they are subjected to constant submergence, a very different experience than those outside of the Baltic Sea. Tides are minimal within the Baltic Sea because the body of water body is so small and the influence from the surrounding tides of the Atlantic and North Sea cannot penetrate into the mostly enclosed sea. The absence of tides is actually quite common in enclosed seas, being seen in both the Black and Caspian seas as well (Medvedev et al., 2016). The absence of tides also plays an important part in stabilising the defined salinity gradient. However if you are on the Baltic Sea coast you might notice that the water level does change from time to time. This is not because of the tide but actually due to air pressure, wind conditions, ice cover and the flow of water between the Baltic and North Sea (Finnish Meteorological Institute, 2017b).
So the Baltic Sea is a semi-enclosed, shallow, brackish sea with various climates, no tides, and a low number of species. As a study system this makes it pretty unique, and adds a further complexity to any research we do in this unique sea. As a marine biologist, I am grateful to have the opportunity to work in such a special place!
Fieldwork at Kõiguste field base (University of Tartu)
Sources:
Alenius, P., Myrberg, K. & Nekrasov, A. 1998. The physical oceanography of the Gulf of Finland: a review. Boreal Environ. Res. 3:97–125.
Eakins, B.W. and G.F. Sharman, Volumes of the World’s Oceans from ETOPO1, NOAA National Geophysical Data Center, Boulder, CO, 2010.
Elmgren, R. & Hill, C. 1997. Ecosystem function at low biodiversity – the Baltic example. Cambridge University Press, Cambridge. 319–336 pp.
Finnish Meteorological Institute, 2017a. Ice season in the Baltic Sea.https://en.ilmatieteenlaitos.fi/ice-season-in-the-baltic-sea [date accessed 28/09/2020]
Finnish Meteorological Institute, 2017b. Sea level variations on the Finnish coast. https://en.ilmatieteenlaitos.fi/sea-level-variations [date accessed 28/09/2020]
Furman, E., Pihlajamäki, M., Välipakka, P. & Myrberg, K. 2014. The Baltic Sea–Environment and Ecology.
Gerlach, S.A. 1994. Oxygen conditions improve when the salinity in the Baltic Sea decreases. Mar. Pollut. Bull. 28:413–6.
HELCOM, 2010a. Ecosystem Health of the Baltic Sea 2003–2007: HELCOM Initial Holistic Assessment. Balt. Sea Environ. Proc. No. 122.
Kuosa, H. & Myrberg, K. 2009. Introduction to the Gulf of Finland Ecosystem. In Rintala, J.-M. & Myrberg, K. [Eds.]. Ministry of the Environment of Finland, Finland, pp. 21–5.
Leppäranta, M. and Myrberg, K., 2009. Physical Oceanography of the Baltic Sea. Springer-Verlag. Berlin-Heidelberg-New York
Medvedev IP, Rabinovich AB and Kulikov EA (2016) Tides in Three Enclosed Basins: The Baltic, Black, and Caspian Seas. Front. Mar. Sci. 3:46. doi: 10.3389/fmars.2016.00046
Seifert, T., Kayser, B. and Tauber, F., 1995. Bathymetry data of the Baltic Sea. Baltic Sea Research Institute, Warnemünde.
Stewart, H.A. and Jamieson, A.J., 2019. The five deeps: The location and depth of the deepest place in each of the world’s oceans. Earth-Science Reviews, 197, p.102896.
Szaniawska, A. 2018. The Gulf of Riga. Springer, Cham.
Waern, M. 1952. Rocky-shore algae in the Öregrund archipelago. Uppsala universitet.
Zillén, L., Conley, D.J., Andrén, T., Andrén, E. & Björck, S. 2008. Past occurrences of hypoxia in the Baltic Sea and the role of climate variability, environmental change and human impact. Earth-Science Rev. 91:77–92.
So in this blog I talk a lot about seaweeds, but what exactly are seaweeds? Seaweed is a common name used to refer to marine macroalgae, a type of large algae. So seaweeds are large algae that live in the sea. But this just seems to raise the bigger question: what are algae?
Most people probably know algae as the clumps of seaweed spoiling the beach or the green stuff that wraps up your sushi.
Seaweed washed up on the beach. Goleta. California. United States.
But algae can come in a vast array of shapes and sizes. The term algae actually encompasses a large range of organisms, from single celled individuals that are a similar size to bacteria, up to large multi-celled, plant-like organisms up to 53m in length in the case of giant kelp.
Coloured electron microscopy of diatoms Arachnoidiscus
Kelp Forest. West coast of North America
Algae are not always alone, and can actually be found forming important symbiotic relationships with many marine animals, the most notable being that with corals, but also with jellyfish, sea anemones and even clams. These algae are provided shelter within the animal whilst in exchange providing nutrients to their animal host in a mutually beneficial relationship.
Algae and Jellyfish symbiosis. Golden Jellyfish Mastigias sp. Jellyfish Lake. Palau
Algae are grouped together in a rather informal way, with many members not necessarily being closely related. These algae can come from multiple lineages, meaning that they are grouped together despite not sharing an immediate common ancestry. This explains why we see such a diverse range of characteristics within the group.
However though a diverse group, they do share one crucial similarity: they are photosynthetic. But then are algae plants? The answer is slightly contentious, but generally, we define them not to be. To be a plant you need a root or internal vascular systems and to produce seeds or flowers, which algae do not. Some algae can have similar looking structures, such as the vein-like midrib in bladderwrack, but these are not true vascular systems. Unlike plants, the body of an alga is relatively undifferentiated, whereby there is no division of labour within the algal body.
Bladderwrack. St. Mary’s Island. NE coast of England.
So algae are a group of photosynthetic organisms that are corralled into a group because they do not fit well into the other more well-defined photosynthetic groups like plants, fungi and lichens. Hence it really isn’t unsurprising that they are so diverse.
The greatest anomaly within algae are the cyanobacteria. Cyanobacteria, also known as blue-green algae, are normally (but not always) included within algae and are probably most famous for forming a greeny sheen of scum on the surface of the water that stops you going in for a swim at the sea or lake side.
Cyanobacteria on the water’s surface
Cyanobacteria are actually very unlike all other algae, even residing within a different kingdom within the tree of life. Cyanobacteria reside within the kingdom of bacteria, alongside the familiar E. coli and Salmonella. Bacteria in general are primitive cells with a less complex structure and are always single-celled. These single cells can exist on their own or as several to many cells living in a colony, chain or filament.
The Cyanobacteria: Oscillatoria and Gleocapsa.
All other algae have a complex cell structure, having many different cellular parts including those specialised for photosynthesis: the chloroplasts. Interestingly, cyanobacteria are considered the origin of chloroplasts, when a cyanobacterial cell was incorporated into the cell of an algal ancestor. Therefore, though cyanobacteria are not directly related to all other algae they are related to the photosynthetic part of them.
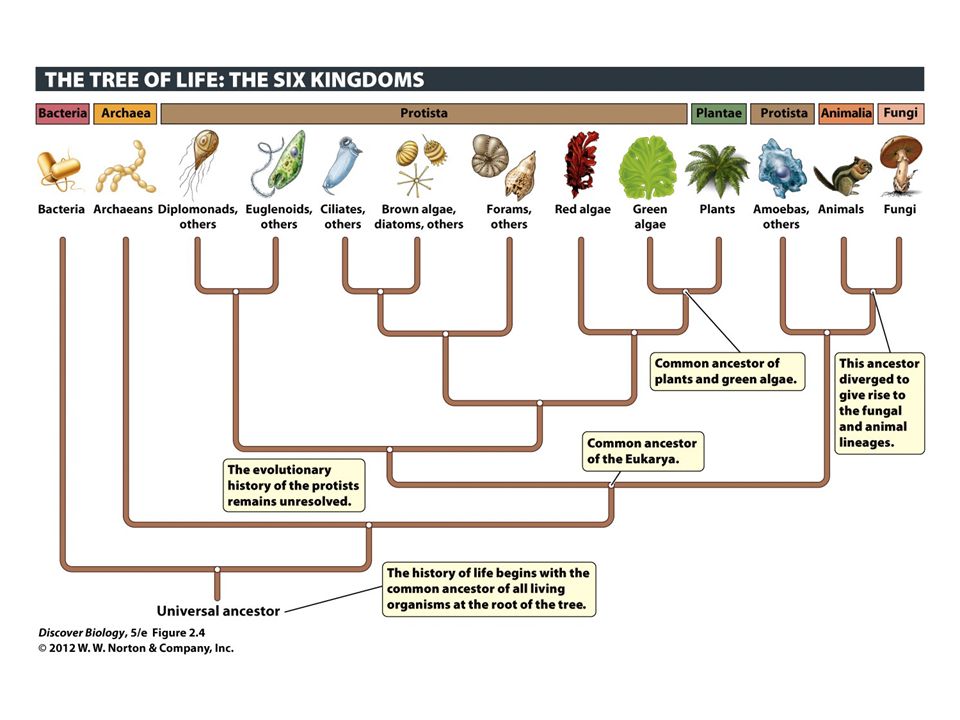
All these algae belong to the kingdom of protista. Of these complex algae, there are three major groups: Green, Red, and Brown. As you may guess they are named for their usual colour.
This evolutionary tree shows the hypothesised relationships among the six kingdoms. Each group branching off the tree can be thought of as a cluster of close relatives. Source: dalconenvironmental.com
The green group contains algal species within the scientific designation Chlorophyta, the red within Rhodophyta, and brown within Heterokonta, Haptophyta, Cryptophyta, and Alveolata. All algae within each of these groups share a set of characteristics that distinguishes them from those within another group, and thus are grouped accordingly.
Red and brown algae. Penguin Island. Western Australia.
The main characteristic difference between these groups are the types of pigment each contains. Just like plants, all these algae contain the green photosynthetic pigment chlorophyll; though the type of chlorophyll varies between them. In green algae we find chlorophyll a and b, which is the same as can be found in plants. Both red and brown algae also contain chlorophyll a, but the second is either chlorophyll d in red algae or chlorophyll c in brown algae. Green, red, and brown algae also contain other different accessory pigments that help chlorophyll in absorbing light. These pigments also contribute to the algae’s colour.
The green algae: Scenedesmus.
There are also a few other smaller groups of algae not contained within these three main groups. These are the freshwater Glaucocystophyta, the predominantly freshwater Euglenophyta, and the marine Chlorarachniophyta. They are all far smaller groups of solely single celled individuals with either 15, ~800 and 5 species respectively. Nevertheless the vast majority of algal species are grouped within the greens, reds and browns.
Green algae are the most closely related algae to plants, sharing a common recent ancestor. They can be found in both marine and freshwater environments. Examples of green algae include sea lettuce (Ulva spp) and Dead man’s fingers (Codium fragile).
Sea lettuce, Ulva spp. Bluefish Point. Manly. Australia
Red algae are mostly marine, and can be found living at greater depths than green and brown algae. This is because they can absorb blue light, which penetrates deeper than other light waves do, and thus they are still able to photosynthesise where other algae cannot.
Some red algae can even be reef builders, similar to that of the corals. Coralline algae are encased within calcareous deposits and form spiky underwater carpets known as maerl beds. These maerl beds are ecologically important, but unfortunately due to their fragility and slow growth are also under threat.
Maerl. Fal and Helford Maerl Beds. UK.
And finally brown algae. As you may guess from the larger number of subgroups included within the main group, brown algae are rather loosely gathered together. Many do not share common recent ancestors. They are mostly marine and also include the largest of the individual algae. Yes that is the kelps, and also bladderwrack and its relatives belong in this group too. But brown algae are not only large, there are also small single celled ones too, including the diatoms. Diatoms are little cells encased in glasshouses, and despite their small size play an important ecological role on a global scale.
The diatom: Coscinodiscus.
So to conclude, algae are the misfits of the tree of life, they do not quite fit anywhere else so are instead lumped together in one big group. There are many differences within the group, but there are also similarities as well. Diversity of algae is vast, from tiny to giant, from simple to complex, but all need light to live. And the number of algae isn’t small either. Currently we have described 50,000 species of algae (Guiry & Guiry, 2020). That’s a lot of algae!
Sources
Guiry, M.D. & Guiry, G.M. 2020. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. https://www.algaebase.org; searched on 28 August 2020.
When asked to picture a tropical marine environment you will probable imagine a tropical coral reef: bright, vivid colours, an abundance of dazzling fish, lovely warm waters and most importantly coral.
Branching coral and resident reef fish. Great Barrier Reef. Cairns. Australia
Yet tropical coral reefs are famously suffering from many threats, both human and environmental. These threats have caused mass coral deaths through the tropical seas. The coral reef cover in the Caribbean has declined from ∼50% in 1977 to ∼10% by 2001 (Gardner et al., 2003). And this phenomenon is not unique, with many other tropical reefs following similar trends.
In the face of these declines there have been shifts from coral dominated reefs to seaweed dominated reefs. In Jamaica, corals declined from more than 50% coverage in the late 1970 to less than 5% in 1994, with fleshy seaweeds coverage increasing to more than 90% at the same time (Hughes, 1994). Indeed, some tropical seaweeds have actually benefitted from coral reef declines caused by coral bleaching, hurricanes, disease and human activities including overfishing and eutrophication. This has allowed tropical seaweeds to increase in abundance, often becoming dominant in what used to be coral dominated reefs.
Bleached coral. Townsville. Queensland. Australia
In fact many scientists and policy makers interpret increases in seaweed as the single best indicator of reef degradation.
But do tropical seaweeds really deserve such a bad reputation?
Tropical seaweeds are often demonised, being credited to the degradation of coral reefs by competition and overgrowth. However tropical seaweeds can in fact play a valuable part in tropical ecosystems as well.
Tropical seaweeds are highly diverse, encompassing many species including larger canopy forming wireweeds (Sargassum) species to smaller, understory lobeweeds (Lobophora) and fanweeds (Dictyota). Many animals rely on tropical seaweeds as food sources or shelter, amongst other things. Various tropical fish, including frogfish and seadragons, can be seen to mimic seaweed as a form of camouflage. In fact the aptly named Sargassum fish, shows incredible resemblance to itsdrifting seaweed home.
Sargassum fish (Histrio histrio). Sargasso Sea. Atlantic Ocean.
Often seaweed reefs can be extensive in comparison to coral reefs. In Ningaloo (Western Australia) seaweed reef coverage was 46% compared to just 8% for coral reefs (Kobryn et al., 2011). Tropical seaweed reefs are especially common in coastal locations and can be considered one of the most prominent types of shallow water tropical habitats. Consequently these tropical reefs can play an important role within interconnected tropical seascapes.
But are all seaweed reefs equal?
Seaweeds have the potential to improve biodiversity, but some seaweed canopies can have far greater effects on diversity than others. Reefs dominated by weedy, low-stature seaweed; often associated with environments pressured by human activity or disturbance; lack the complexity required to support a plethora of plants and animals. On the other hand, seaweed reefs containing a mixture of canopy and understorey seaweeds and coral provide a high-complexity habitat that can support many levels of biodiversity.
Thus habitat quality; encompassing the coverage, density and canopy height; is important in influencing the range of plants and animals that a seaweed reef can support.
Fish recruits, including important fishery target species; have a much better survival in seaweed reefs with high quality canopies. Therefore more of these new fish recruits can join the adult populations in both coral and seaweed reefs. Consequently seaweed reefs with high quality canopies are much better at supporting fish populations, both within the seaweed reef and outside of it, compared to those with low quality.
Figure 3(b) from Fulton et al., 2019. The image show the influence that canopy quality has on both juvenile and adult reef fish abundance. High quality canopies provide a more complex habitat for juvenile reef fish (black). This increases the survival of juvenile reef fish that can then replenish adult populations in both coral (grey) and seaweed (blue) reefs.
So tropical reefs can, in certain circumstances, be beneficial despite their bad reputation.
Unfortunately, their commonness in coastal areas and the belief that they are best indicators for reef degradation has increased the risk that seaweed reefs are erroneously concluded to be a result of detrimental human activity rather than a natural component of a healthy ecosystem.
In seaweed reefs, if there is no evidence that a shift from coral to seaweed has occurred then they should be considered natural components of the seascape.
Brown Fan Weed (Dictyota sp.). Bare Island. New South Wales. Australia
In fact seaweed reefs can often be found on coastlines and remote reefs with low pressure from human activity, being an important part of the interconnected mosaic of tropical patch habitats. These reefs merit protection and deserve recognition as key components of the tropical seascape.
So not all seaweed reefs are the same: some are indicators of an unhealthy coral reef system though others are natural tropical habitats. We need to acknowledge the assets that complex seaweed reefs can be and protect them.
This post was based on the research of Fulton et al., (2019).
If you enjoyed this post you can find the article here:
Gardner, T.A., Côté, I.M., Gill, J.A., Grant, A. and Watkinson, A.R., 2003. Long-term region-wide declines in Caribbean corals. science, 301(5635), pp.958-960.
Hughes, T.P., 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science, 265(5178), pp.1547-1551.
Kobryn, H.T., Wouters, K. and Beckley, L.E., 2011. Ningaloo Collaboration Cluster: Habitats of the Ningaloo Reef and adjacent coastal areas determined through hyperspectral imagery.
The definition of a species states that all individuals within a species will have the same main characteristics (Collins Dictionary, 2020). These Burchell’s zebras all share the same characteristic striped appearance for example.
A group of Burchell’s zebras. Maasai Mara National Reserve, Kenya
So each individual within a species must share similar characteristics. But does this mean each will respond to environmental change in the same way? Traditionally a species is assumed to be one homogenous unit, assuming that all individuals within the species have similar environmental tolerances irrespective of their origin within the species range. Accordingly all individuals should be able to exist anywhere within the species’ range.
But is it that simple? Do all individuals respond the same way or do some individuals have the ability to respond differently? And more pressingly: Will all individuals within a species be affected by climate change in the same way?
Coal Powerplant; Kohlekraftwerk. Lünen, Germany
Greenhouse gas emissions caused by human activity have enhanced the greenhouse effect; resulting in additional warming of the Earth’s surface (Houghton et al., 1990). Of this extra heat, 93% has been absorbed by the ocean leading to surface water warming of 0.11°C per decade in the Indian Ocean, 0.07°C per decade in the Atlantic and 0.05°C per decade in the Pacific (Hoegh-Guldberg et al., 2014).
Increased mean temperatures are redistributing species across the globe with many cold-water species moving polewards; including brown seaweeds. In Europe the cold-water cuvie kelp is already showing range shifts toward northern regions and local extinctions in its southern range (Müller et al., 2009) and by 2200 two common rockweeds will be lost from European shores south of Bordeaux (Jueterbock et al., 2013).
Forest kelp or cuvie (Laminaria hyperborea). Shetland, UK
If all individuals of a species respond the same way to warming then those at the edge of the species’ range will be more likely to be subjected to temperatures outside of their tolerance. These edge populations will therefore be most susceptible to range shifts and local extinctions. Those individuals within the central range should have tolerance to the warming since there is a greater margin before they are exposed to intolerable temperatures. Thus the species should be maintained within the central range and lost at the edges.
But what if all individuals do not respond the same way? In that case central populations could be in just as vulnerable a state.
If individuals respond differently, than those from different locations might be able to tolerate temperature changes to different degrees. Therefore those in central populations may also be exposed to temperatures outside of their tolerance.
Consequently if all individuals do not respond to warming in the same way then the species could be vulnerable to climate change throughout its whole range.
So either all individuals are the same and the edge populations are most vulnerable or individuals can be different and then all populations could be vulnerable.
Gulfweed (Sargassum), Rockweed (Fucus) and Sea lettuce (Ulva). Llangennith, Wales, United Kingdom
Well, a systematic review of literature relating to marine plants and seaweeds appears to help answer this question. A striking 90% of studies observed within species differences to temperature change, meaning that individuals do in fact respond differently to temperature (King et al., 2018). Therefore all populations could be vulnerable. We could see local extinctions from both the edge and the central range of many important seaweed species.
To add to that, many seaweeds have limited dispersal. This results in highly structured distributions with closely related individuals remaining in close proximity. These individuals of close relation will possess similar characteristics, including temperature tolerance. Accordingly most individuals within the population will respond similarly to warming.
Sardine run. Tonga
In a highly mobile species with high dispersal, for example many fish species, migrant fish can move polewards to track the tolerable temperature and replace previously resident fish that can no longer tolerate the temperature in their previous habitat. These migrants can also contribute vital genetic material to the populations allowing the population to adapt to warming.
But seaweeds are not mobile; they cannot migrate since they are generally permanently attached to a hard surface. Consequently migration cannot readily contribute to the population. This means that struggling resident seaweeds cannot be replaced by migrant seaweeds. No migration and limited dispersal also means that no new genetic material is added to the population, therefore hindering adaptation to warming.
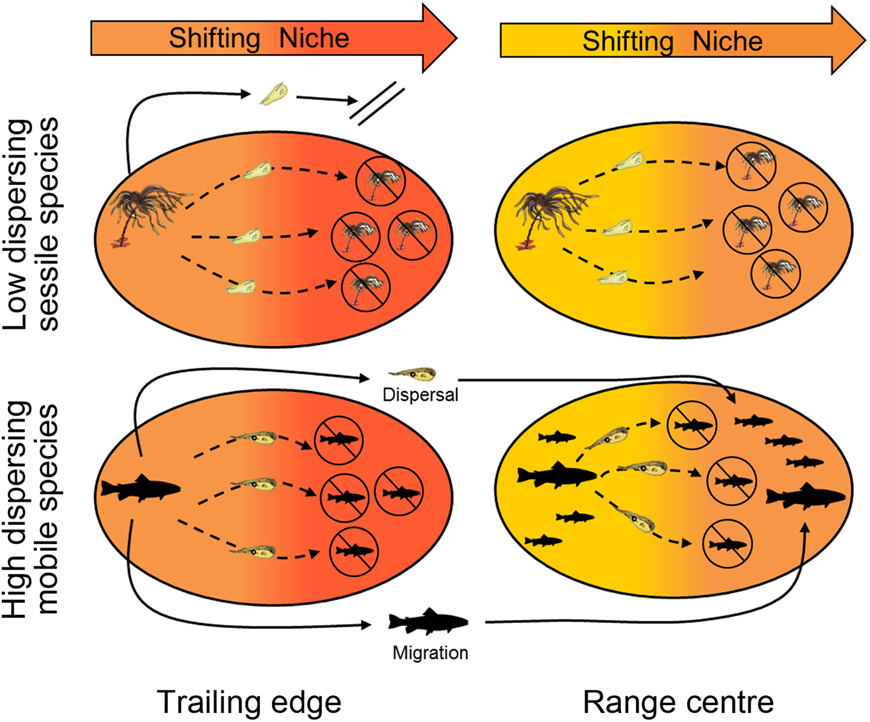
Figure 1 from King et al., 2018 Diagrammatic representation of how low and high dispersing species will respond to warming at the edge and centre of their range. Ovals represent the experienced warming within the edge and central range. Under warming conditions low dispersing species, such as seaweeds, with individuals that tolerate different temperatures will not be able to disperse into future suitable habitats and will become locally extinct.
It seems then that whether a population is at the edge or in the centre of the species range warming can be disastrous for seaweeds.
But individuals having different tolerances might also help protect a species, given a human little help. If individuals tolerant to warming are relocated to population susceptible to warming we may be able to boost local temperature tolerances. This concept of ‘Assisted migration’ is not new, having already been developed in terrestrial forests (McLachlan et al., 2007; Aitken & Whitlock 2013; Williams and Dumroese, 2013) and seagrass meadows (Katwijk et al., 2016).
There are far more inherent logistical problems relocating seaweeds compared to the far simpler reseeding of terrestrial trees and seagrasses, however there is growing indication that this may become a feasible option in the face of climate change.
So we can conclude that just because all individuals from a species must share similar characteristics this does not mean that they are all exactly the same. We need to stop considering species as a single homogenous unit and instead see all individuals as unique in their own small way.
This post was based off the research of King et al., (2018).
If you enjoyed this post you can find the article here:
Aitken, S. N. and Whitlock, M. C. 2013. Assisted gene flow to facilitate local adaptation to climate change. Annual Review of Ecology, Evolution, and Systematics. 44: 367–388.
Houghton, J.T., Jenkins, G.J. and Ephraums, J.J., (1990) Climate change: the IPCC scientific assessment. Intergovernmental Panel on Climate change (IPCC), Cambridge University Press; Cambridge, United Kingdom
Hoegh-Guldberg, O., R. Cai, E.S. Poloczanska, P.G. Brewer, S. Sundby, K. Hilmi, V.J. Fabry, and S. Jung, 2014: The Ocean. In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Barros, V.R., C.B. Field, D.J. Dokken, M.D. Mastrandrea, K.J. Mach, T.E. Bilir, M. Chatterjee, K.L. Ebi, Y.O. Estrada, R.C. Genova, B. Girma, E.S. Kissel, A.N. Levy, S. MacCracken, P.R. Mastrandrea, and L.L.White (eds.)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 1655-1731.
Katwijk, M. M. et al. 2016. Global analysis of seagrass restoration: the importance of large-scale planting. Journal of Applied Ecology. 53: 567–578.
King, N.G., McKeown, N.J., Smale, D.A. and Moore, P.J., 2018. The importance of phenotypic plasticity and local adaptation in driving intraspecific variability in thermal niches of marine macrophytes. Ecography, 41(9): 1469-1484.
McLachlan, J. S. et al. 2007. A framework for debate of assisted migration in an era of climate change. Conservation Biology 21: 297–302.
Müller, R., Laepple, T., Bartsch, I. and Wiencke, C., 2009. Impact of oceanic warming on the distribution of seaweeds in polar and cold-temperate waters. Botanica Marina, 52(6): 617-638.
Jueterbock, A., Tyberghein, L., Verbruggen, H., Coyer, J.A., Olsen, J.L. and Hoarau, G., 2013. Climate change impact on seaweed meadow distribution in the North Atlantic rocky intertidal. Ecology and evolution, 3(5):1356-1373.
Williams, M. I. and Dumroese, R. K. 2013. Growing assisted migration: synthesis of a climate change adaptation strategy. – In: Haase, D. L. et al. (tech. coord.), National Proceedings: Forest and Conservation Nursery Associations – 2012. Proceedings RMRS-P-69. Fort Collins, CO, USA Dept of Agriculture, Forest Service, Rocky Mountain Research Station, 90–96.
The ice is disappearing… Over this winter there was no walking on the sea around Helsinki, or in much of Finland for that matter.
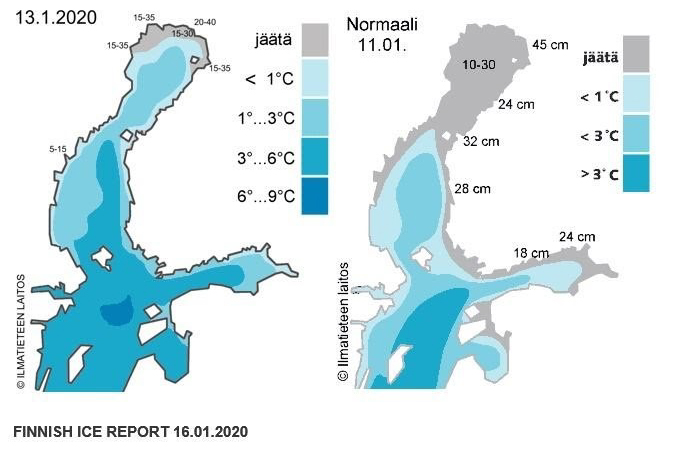
In January the ice cover was dramatically lower than normal for the Baltic Sea. Normally the ice should extend from the northern parts of the Baltic down to the Gulf of Finland and the Gulf of Riga, however this year the ice was limited to just a small section of the Bothnian Bay.
Indeed this lack of ice is a tell-tale sign of a milder winter, both on land and in the sea.
Extent of ice coverage (jäätä) in the northern Baltic Sea under normal condition and those observed in January 2020. Number on ice indicate thickness. Source: Finnish Meteorological Institute
But was last winter an odd exception?
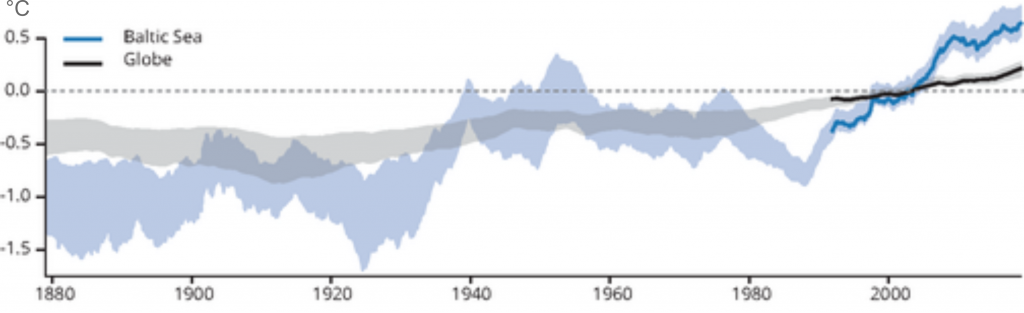
It appears not. The earth is experiencing a warming of the marine environment in general, but the Baltic Sea is warming far more severely than the average global ocean. The Baltic Sea temperature is now higher than that average temperature recorded between 1981-2010 (European Environmental Agency, 2019)
Decadal global and Baltic Sea averaged sea-surface temperature anomalies relative to a 1981-2010 baseline. Source: European environmental agency
However it is not solely the increase in temperature that is concerning for the marine environment. Shorter spells of prolonged warmer temperatures, called marine heatwaves, are also troubling.
Due to climate change the IPCC (2013) expects marine heatwaves to become more frequent, and those that we do experience to be far more intense. Already the marine environment has experienced a 54% increase in annual marine heatwave days (Oliver et al., 2018).
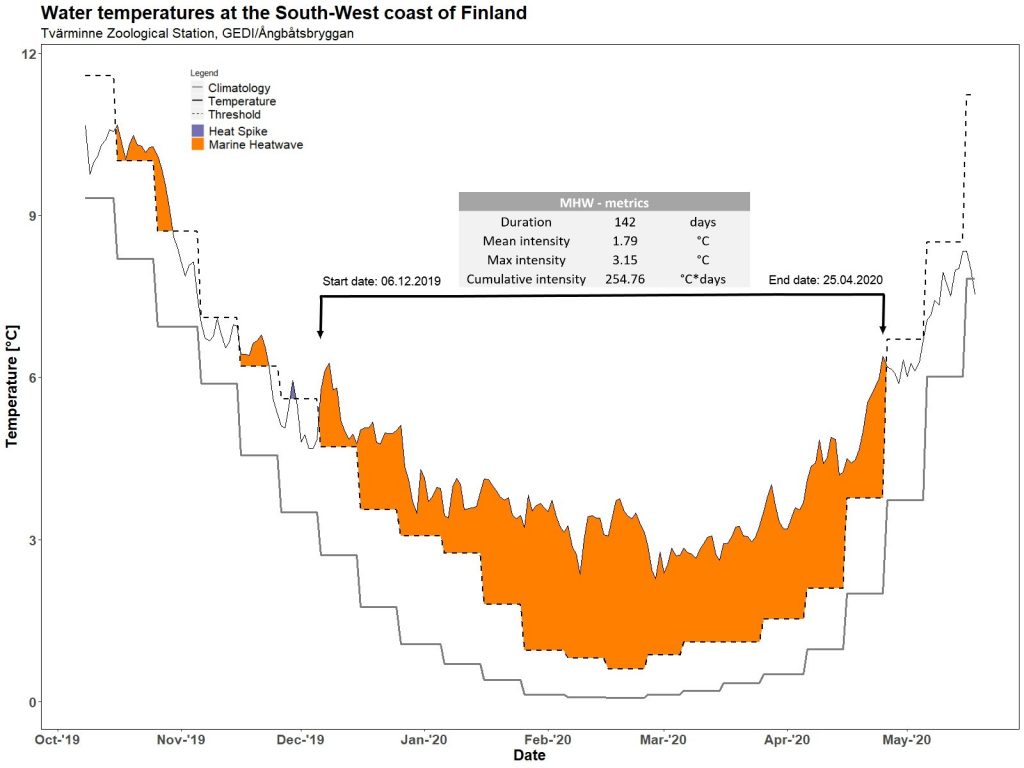
And Finland is no exception. Monitoring at the University of Helsinki’s Tvärminne Zoological Station points that last winter was just one long marine heatwave. Exceptionally warm conditions lasted an outstanding 142 days, and even included record breaking temperatures in February 2020 (@Tvärminne, 2020).
Water temperature over the winter of 2019/2020 recorded by temperature loggers in shallow areas near Tvärminne. Source: Tvärminne Zoological Station .
Are marine heatwaves really all that bad?
Prolonged high temperatures can induce heat stress in marine plants and seaweeds with disruptive consequences to the survival of these organisms.
This stress can reduce a marine plant or algae’s ability to persist within previously inhabited areas. Consequently heatwaves have the potential to cause local extinctions.
By 2200 three currently common seaweeds; bladder, serrated and knotted wrack; will have disappeared from any North-Atlantic shore south of 45 ° latitude due to marine heatwaves and the other effects of climate change (Jueterbock et al., 2013).
How will marine heatwaves affect Baltic Sea ecosystems?
Recent research at GEOMAR Helmholtz Centre for Ocean Research sheds some light on this.
By using large, outdoor tanks called ‘mesocosms’, researchers tested how eelgrass and bladderwrack respond to heatwaves (Saha et al., 2019). Within these 1,500l tanks the eelgrass and bladderwrack were subjected to 9 day heatwaves.
Well is seems that both eelgrass and bladderwrack are fairly resilient to heatwaves. The eelgrass did experience impaired growth, but also showed good signs for potential recovery. Bladderwrack, on the other hand, showed no impaired growth with the only significant effect being seen on the bacterial community living on the algae.
Therefore it seems that both eelgrass and bladderwrack will likely be able to endure short heatwaves similar to those of this experiment when experienced within the natural environment.
However it is not all good news. The effects of long heatwaves will probably be fairly different. In a previous study subjecting eelgrass to a continuous 3 week heatwave far greater detrimental effects where observed (Winters et al., 2011).
So a tolerance to short heatwaves does not necessarily equate to a tolerance to longer ones. Consequently the length of each heatwave will be critical to determining the effects on eelgrass and bladderwrack.
We will have to endure more and hotter marine heatwaves, as will the resident plants and animals of the Baltic Sea, yet at least eelgrass and bladderwrack seem somewhat prepared for the future.
European Environmental Agency, 2019. Decadal average sea surface temperature anomaly in different European seas. Date accessed: 29/5/20. https://www.eea.europa.eu/legal/copyright
IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Stocker, T.F., D. Qin, G.-K. Plattner, M. Tignor, S.K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex and P.M. Midgley (eds.)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 1535 pp, doi:10.1017/CBO9781107415324.
Jueterbock, A., Tyberghein, L., Verbruggen, H., Coyer, J.A., Olsen, J.L. and Hoarau, G., 2013. Climate change impact on seaweed meadow distribution in the North Atlantic rocky intertidal. Ecology and evolution, 3(5), pp.1356-1373.
Saha, M., Barboza, F.R., Somerfield, P.J., Al‐Janabi, B., Beck, M., Brakel, J., Ito, M., Pansch, C., Nascimento‐Schulze, J.C., Jakobsson Thor, S. and Weinberger, F., 2020. Response of foundation macrophytes to near‐natural simulated marine heatwaves. Global Change Biology, 26(2), pp.417-430.
Winters, G., Nelle, P., Fricke, B., Rauch, G. and Reusch, T.B., 2011. Effects of a simulated heat wave on photophysiology and gene expression of high-and low-latitude populations of Zostera marina. Marine Ecology Progress Series, 435, pp.83-95.
We are currently in a biodiversity crisis. The rate of extinction is 100 to 1000 times faster than the natural rate (Singh, 2012) and by the end of the 21st century we will have lost at least 50% of all species (Myers, 1993).
A view from space
Undoubtedly, this species loss will have overwhelming effects on how we live. Biodiversity loss is not only an environmental issue, it also directly effects us as a society, as well as our development, economy and security. In fact the UN (2019) declared that biodiversity loss undermines progress towards 80% of the assessed targets of their Sustainable Development Goals.
Much attention has been focused on the loss of species, such as the charismatic black rhino or the snow leopard. With more than 31,000 species threatened with extinction (IUCN, 2020) this is understandable. Species are undeniably important and we should aim to conserve them, but we should not ignore the fact that we are losing so much more than just species in this biodiversity decline.
Snow leopard, Marwell Zoo, UK
Biodiversity manifests throughout scales, from ecosystems, communities, species, and even genes. Genetic diversity, or the variety of genes, is a vital part of biodiversity. Yet genetic diversity is frequently overlooked in international conservation policies.
Genetic diversity can be represented between species: you are genetically different from your pet cat; or within species: both you and I share different genes. If genetic diversity declines within a species there can be devastating consequences to the stability and functioning of that species.
In the case of the Florida panther low genetic diversity led to a range of severe problems. In fact the species was slated for extinction, with only ~30 individuals remaining (Ohio State University, 2019). Luckily for the Florida panther this downward spiral was rescued by the addition of new genes into the genepool from eight Texas puma females. This increased genetic diversity has saved the Florida panther and now at least 230 roam the wilds of Florida.
But what about species that have natural populations with sustained low genetic diversity?
Seagrass meadow, Rottnest Island, Western Australia
Seagrasses have notoriously low genetic diversity, often forming vast meadows of a few clones. The genetic diversity within these meadows, though conventionally low, is also important. Seagrass meadows with greater numbers of genetically different individuals have far better growth after disturbance compared to meadows of a single clone (Reusch et al., 2005; Hughes & Stachowicz, 2011). So even in species with typically low genetic diversity, higher diversity can be beneficial.
This highlights that policy makers are making a big mistake overlooking genetic diversity. By taking genetic diversity into account we could vastly improve the effectiveness of our conservation measures. Hence, if we want to successfully conserve a species, especially a threatened species, the conservation of the genetic variation within the species is of the utmost importance.
Sources:
Hughes AR, Stachowicz JJ. 2011. Seagrass genotypic diversity increases disturbance response via complementarity and dominance. Journal of Ecology 99:445-453
IUCN 2020. The IUCN Red List of Threatened Species. Version 2020-1. https://www.iucnredlist.org. Downloaded 30 April 2020
Myers, N. (1993). Biodiversity and the precautionary principle. Ambio, 22, 2/3:74-79
Ohio State University. (2019, October 3). How the Texas puma saved the Florida panther: Uncovering the genetic details of a conservation success story. ScienceDaily. www.sciencedaily.com/releases/2019/10/191003111755.htm. Downloaded 29 April 2020
Reusch TBH, Ehlers A, Hämmerli A, Worm B. 2005. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proceedings of the National Academy of Sciences of the United States of America 102:2826_2831 DOI 10.1073/pnas.0500008102
Singh, J. S. (2002). The biodiversity crisis: a multifaceted review. Current Science, 82(6), 638-647
Beyond the range of conventional diving and above where
submersibles normally roam lies a unique habit rarely studied. Exploration of
this new world leads to the discovery of a new wealth of biodiversity; upon
each exhibition to the deep scientists come back with brand new species never
known to science before.
But does seaweed really live 100m below the surface where light barely touches?
NOAA scientific divers descending to 150ft, Pearl and Hermes Atoll, Hawaii
Well yes they do, in fact they can be even more abundant
than the neighbouring corals. Carpets of reds and greens, with shoots up to 1m long,
bloom throughout this range. Not only that but walls of mushroom-like brown
algae and beds of fine green seaweed are amongst the other seaweeds that can be
found.
So even in deep waters seaweeds are important players in
the marine ecosystem.
And if you ever wondered what it entails to be a seaweed researcher then this is a great example. Here at FunkVeg we use many of the same techniques and can vouch for the completely new level of awkwardness these pesky critters create.
What do kelp forests, mussel beds, seagrass meadows and coral reefs have in common? They are all given their name by the foundation species that creates them.
But what does being a foundation species entail?
Great Barrier Reef, Australia
Just as the foundations of your house provide structural stability keeping you house strong and stable, so too does a foundation species within an ecosystem. Because of this they are vitally important in structuring the community and maintaining a healthy ecosystem.
With
the growing influence and uncertainty of human activity and environmental
change the stability of the world’s ecosystems is questionable. It is therefore
imperative that we understand what keeps ecosystems stable.
‘Does a stable giant kelp forest result in a more stable understory community?´
Anemone in Kelp, Monterey Bay National Marine Sanctuary, USA
Well it turns out it does. Just as stable foundations ensure your house stays up, so too does a stable forest of kelp help in maintaining the stability of the under-story community of plants and animal.
Unfortunately in the face of climate change the stability of kelp is likely to change in the future. This could have devastating effects of all the plants and animals that rely on the kelp forest. Not only this, but this new research suggests that all habitats reliant on foundations species may also face such similar problems. Without the important species providing a foundation for all the other plants and animals, we could see devastating changes to these iconic habitats.
So
the take home note:
Just as you have to take care of the foundations of your house to prevent it collapsing, so too do we have to take care of the foundations of our ecosystems or face calamitous consequences.
Scorpionfish in Seagrass, NOAA Florida Keys National Marine Sanctuary, USA
If you want to read more about this research check out this link or article below:
Lamy, T., Koenigs, C., Holbrook, S.J., Miller, R.J., Stier, A.C. and Reed, D.C., 2020. Foundation species promote community stability by increasing diversity in a giant kelp forest. Ecology, p.e02987.