Oppimateriaali ”Nisäkkäät” on suunniteltu 7. luokan biologian opetukseen.
Oppimateriaali pdf-muodossa:
BIO-004 oppimateriaali NISÄKKÄÄT
Oppimateriaalin ovat toteuttaneet Nelli Aalto, Lauri Perkiö ja Susanna Stenberg.
Oppimateriaali ”Nisäkkäät” on suunniteltu 7. luokan biologian opetukseen.
Oppimateriaali pdf-muodossa:
BIO-004 oppimateriaali NISÄKKÄÄT
Oppimateriaalin ovat toteuttaneet Nelli Aalto, Lauri Perkiö ja Susanna Stenberg.
Tekijät: Senni Luoto, Sonja Pekkola ja Laura Ahola
Tämä oppimateriaali on tuotettu Johdatus biologian opetukseen -kurssin puitteissa AAKE-hankkeeseen. Teoriaosuuden lisäksi postaukseen kuuluu tehtäväpaketti sekä mallivastaukset.
Eliökunnan historia voidaan jakaa neljään pääaikakauteen: elämän alkuaikaan, elämän vanhaan aikaan, elämän keskiaikaan ja elämän uutteen aikaan. Elämän alkuajalla eli prekambrisella ajalla ilmakehässä oli hyvin vähän happea ja eliökunta koostui pääasiassa merien arkeista, bakteereista ja syanobakteereista. Elämän alkuaika päättyi n. 570 miljoonaa vuotta sitten niin kutsuttuun kambrin räjähdykseen. Kambrin räjähdyksellä viitataan siihen, että eliömäärä lisääntyi merissä räjähdysmäisesti. Tämä oli alkusysäys seuraavalle aikakaudelle – elämän vanhalle ajalle eli paleotsooiselle ajalle.
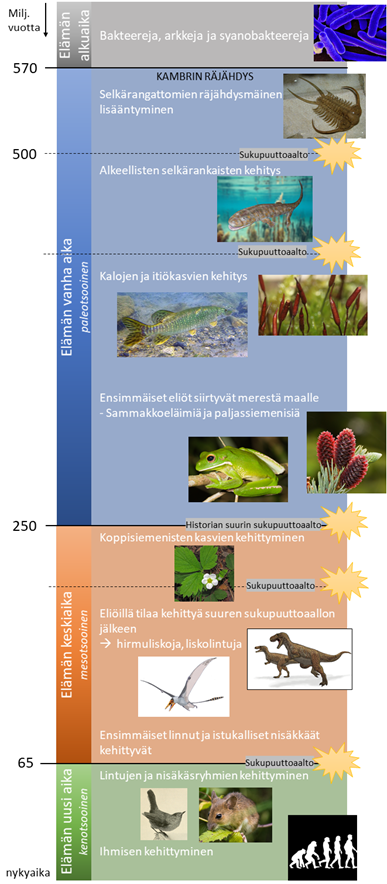
Kaaviokuva: Senni Luoto, kuvat: Creative Commons
Elämän vanhan ajan alussa merien eläimille alkoi kehittyä alkeellista hermostoa ja näköaistia, minkä ansiosta myös ensimmäiset petoeläimet kehittyivät. Myös ensimmäiset nykypäivän merkittävät fossiilit syntyivät, kuten runsaslukuiset trilobiitit. Vanhan ajan loppupuolella elämä siirtyi asteittain merestä maalle, mikä vaati eliöiltä uudenlaisia sopeumia. Elämän historian aikakausiin merkittävästi vaikuttaneita tapahtumia oli suuret sukupuuttoaallot, joita tapahtui elämän vanhan ajan aikana kolme. Viimeisin näistä vanhan ajan sukupuuttoaalloista n. 250 miljoonaa vuotta sitten on maapallon historian tuhoisin. Äkillinen lämpötilan nousu hävitti maapallolta kaikkiaan n. 96% lajeista. Tästä sukupuuttoaallosta alkoi uusi aikakausi – elämän keskiaika eli mesotsooinen aika.
Elämän keskiajan alussa elintilaa valtavan massasukupuuton jälkeen oli paljon. Ilmasto oli kuuma, mistä todisteena ovat pohjoisilta vuoristoilta löytyneet sanikkaisfossiilit. Elämän keskiaika oli dinosaurusten ja liskolintujen sekä suurien koppisiemenisten valtakautta. Tämä valtakausi päättyi kuitenkin n. 65 miljoonaa vuotta sitten asteroidin iskeytyessä Meksikoon Jukatanin niemimaan alueelle, synnyttäen valtaisan tsunamin ja nostattaen pölypilven, joka esti auringonsäteiden pääsyn maapallolle. Näitä tapahtumia seurasi pitkä kylmä jakso, jonka aikana lähes 75% maapallon eliöistä kuoli sukupuuttoon – mukaan lukien dinosaurukset.
Maapallolla on siis tapahtunut kaikkiaan 5 suurta sukupuuttoaaltoa. Niistä viimeisin käynnisti samalla nykyisen aikamme. Elämän uusi aika eli kenotsooinen aika on nisäkkäiden evoluution kulta-aikaa, joka lopulta johti myös ihmisen kehittymiseen. Ihmisen toiminnan myötä maapallo on muuttunut paljon ja monia lajeja on kuollut sukupuuttoon. Siksi yleinen käsitys onkin, että tällä hetkellä on käynnissä kuudes, ihmisen aiheuttama sukupuuttoaalto, jonka seurauksena vuoteen 2020 mennessä selkärankaisista lajeista jopa kaksi kolmasosaa saattaa olla tuhoutunut.
Nykyisin arvellaan, että elämä maapallolla saattoi hyvinkin syntyä syvänmeren alueella. Tuolla, katseilta piilossa, on maailman suurin geologinen muodostuma. Se on valtamerten keskiosia peittävä vedenalainen vuoristo, joka ulottuu maapallon kaikkiin osiin. Tätä valtavaa vuoristo kutsutaan keskiselänteeksi. Vilkaise kuvaa 2 ja kuvittele, kuinka eteläisellä Tyynellämerellä merenpohja repeilee. Yli kolme neljäsosaa planeettamme tuliperäisestä toiminnasta tapahtuu syvänmeren alueella. Toiminta on keskittynyt valtamerten keskiselänteille. Rajuista purkauksista seuraa kuitenkin paljon hyvää. Halkeamista syöksyy kaasuja ja tulikuuma vettä. Näistä suihkuista tiivistyvät mineraalit muodostavat suuria piippuja, joita kutsutaan hydrotermisiksi purkausaukoiksi. Eräät arkeonit ja bakteerit viihtyvät näissä aukoissa. Nykyisin tiedetään myös, että näiden kuumien lähteiden ympärillä on yhtä paljon elämää kuin trooppisissa sademetsissä. Yhden neliömetrin alalle voi paikoitellen ahtautua jopa puoli miljoonaa yksittäistä eläintä.
Hydrotermisistä purkausaukoista ehkä tärkeimmät sijaitsevat Atlantilla. Tämä erityinen purkausaukkojen alue on nimetty kadonneeksi kaupungiksi. Sen 60 metristen tornien sisällä tapahtuu jotain todella poikkeuksellista. Äärimmäisessä paineessa ja lämpötilassa syntyy spontaanisti hiilivetyjä, molekyylejä, jotka ovat kaiken elollisen peruskomponentteja (orgaanisen kemian perusta). Monet tutkijat uskovat nykyään, että elämä maapallolla saattoi saada alkunsa tällaisen purkausaukon ympärillä 4 miljardia vuotta sitten. Jos elämää esiintyy näin äärimmäisissä oloissa maapallolla, sitä voi varmasti esiintyä myös muualla.

Hydroterminen purkasaaukko eli syvänmeren savuttaja
Credit: New Zealand-American Submarine Ring of Fire 2005

Merenalainen vulkaaninen purkaus Tyynellämerellä,
Credit: Image courtesy of Submarine Ring of Fire 2006 Exploration
Elämän alkuaikana ensimmäiset monisoluiset eliöt kehittyivät yhdistymällä monista yksisoluisista. Esitumalliset kehittyivät siis aitotumallisiksi eliöiksi, joilla ei vielä alkuun ollut tuman lisäksi muita soluelimiä. Ajan kuluessa monisoluisuus mahdollisti solujen erikoistumisen omiin tehtäviinsä, sekä eliön tehokkaamman kasvun ja suuremman koon. Suvuttoman lisääntymisen rinnalle kehittyi myös uusi, tehokkaampi keino lisääntyä. Suvullinen lisääntyminen mahdollisti perinnöllisen muuntelun sekä nopeamman sopeutumisen muuttuvan ympäristön uusiin olosuhteisiin. Kemo- ja fotosynteesin myötä eliöt pystyivät myös tuottamaan itse oman energiansa. Ensimmäisiä fotosynteesiin eli valon avulla yhteyttämiseen kykenevät eliöt olivat merissä eläviä syanobakteereita. Yhteyttämisen sivutuotteesta hapesta muodostui vähitellen ilmakehään UV-säteiltä suojaava otsonikerros. Vaikka aluksi happi oli anaerobisiin olosuhteisiin tottuneille eliöille myrkyllistä, soluhengityksen kehittyessä ja sopeutumislevittäytymisen kiihtyessä elämä alkoi siirtyä hiljalleen merestä maalle.
Kambrikauden räjähdyksen myötä elämän vanhalla ajalla kehittyivät eliöiden kaikkien nykyisten pääjaksojen edustajat. Vaikka suurin osa eliöistä eli yhä merissä, kilpailu maan valloittamisesta oli alkanut. Endosymbioositeorian mukaan viherhiukkaset ovat ilmestyneet soluun sen fagosytoimista bakteereista. Yhteyttävistä viherlevistä kehittyivät ensimmäiset maakasvit eli sammalet, sekä myöhemmin sanikkaiset ja paljassiemeniset siemenkasvit. Ensimmäiset maaeläimet puolestaan olivat selkärangattomia kuten niveljalkaisia, jotka sopeuduttuaan aerobisiin oloihin saivat kilpailuedut siirtyessään ravinnon perässä maalle. Merten ensimmäisistä selkärankaisista eli luu- ja rustokaloista kehittyi hiljalleen sammakkoeläimiä, jotka puolestaan olivat ensimmäisiä maalla eläviä selkärangallisia. Sammakkoeläimistä kehittyi ajan myötä matelijoita ja niistä edelleen ensimmäisiä lintuja ja nisäkkäitä.

Pölyttäjän ja kukkakasvin rinnakkaisevoluutio on yksi esimerkki koevoluutiosta.
Kuva: Public Domain.
Koevoluution eli rinnakkaisevoluution myötä elämän keskiajalla kehittyivät myös koppisiemeniset kukkakasvit sekä niitä pölyttävät hyönteiset. Sekä kasvien että eläinten tuli sopeutua lyhyessä ajassa suuriin muutoksiin. Uudet kehittyvät rakenteet mahdollistivat uusien lajien synnyn ja sopeutumisen maaelämään. Permikauden lopulla uusi massasukupuutto katkaisi matelijoiden valtakauden ja linnut ja nisäkkäät alkoivat kehittyä ja runsastua edelleen. Lintujen ja nisäkkäiden, sekä uusien kasvien evoluutio jatkui voimakkaasti elämän uudella ajalla, jonka lopulla kehittyi myös nykyajan ihminen.
2.
Tämä oppimateriaali on tuotettu osana Helsingin yliopiston biologian aineenopettajaopintojen Johdatus biologian opiskeluun -kurssia. Perehdyimme entsyymeihin ja kirjoitimme tietotekstin omaa ymmärrystämme syventäen. Suunnittelimme ja toteutimme myös aiheeseen liittyvän opetustuokion lukion biologian Solu ja perinnöllisyys -kurssin (BI3) opiskelijoille.
Tekijät: Katja Leiman, Henna Helén ja Sonja Vanhanen
Kaikki elävät eliöt tarvitsevat entsyymejä toimiakseen (12). Ilman entsyymejä elämälle välttämättömät reaktiot tapahtuisivat liian hitaasti, ja onkin esitetty, että ilman niitä elämä ei olisi mahdollista. (17, s. 29.) Esimerkiksi rubisco helpottaa hiilidioksidin sitomista ilmasta kasvien fotosynteesin pimeäreaktiossa, ja sitä onkin sanottu maailman tärkeimmäksi entsyymiksi (13, s. 19). Myös ihmisen elimistön kemiallisista reaktioista useimmat ovat entsymaattisia (17, s. 29), ja entsyymit ovat äärimmäisen tärkeässä roolissa mm. ruoansulatuksessa (12). Ilman ruuansulatusentsyymejä eläimet ja ihmiset kuolisivat nälkään, sillä entsyymit nopeuttavat ruuan pilkkoutumista solujen käyttöön sopivaan muotoon (7).
Entsyymejä on jokaisessa solussa ja tuhansia erilaisia (1 s. 28; 20, s. 54). Suurin osa niistä toimii solun sisällä, mutta osa eritetään myös solun ulkopuolelle (18). Solun sisäpuolella toimivia entsyymejä ovat esimerkiksi DNA:n kaksoiskierteen avaava entsyymi RNA-polymeraasi sekä lysosomissa ravintoaineita ja nukleiinihappoja pilkkova hydrolaasi (13). Solun ulkopuolelle erittyviä entsyymejä taas ovat esimerkiksi ruuansulatusentsyymit (18).
Kaikki solut tuottavat entsyymejä (18), ja niitä valmistetaan ja eritetään elimistössä tarpeen mukaan (13). Solun tumassa olevat geenit ohjaavat, millaisia entsyymejä ja kuinka paljon solu valmistaa (13).
Useimmat entsyymit ovat proteiineja (25). Solut valmistavat proteiineja aminohapoista proteiinisynteesin avulla. Aminohappojen muodostamat aminohappoketjut kiertyvät tai laskostuvat, vääntyvät mutkille ja liittyvät yhteen pallomaisiksi entsyymeiksi monivaiheisen prosessin seurauksena. (13.) Aminohappojärjestys määrää entsyymin kolmiulotteisen rakenteen (7). Tietyt entsyymit vastaavat oikeanlaisen kolmiulotteisen rakenteen muodostumisesta (19, s. 138). Entsyymin rakenne puolestaan määrittää sen, mitä kemiallista reaktiota entsyymi katalysoi (7). Entsyymi voi myös olla virheellinen, eikä sitä aina valmistu ollenkaan. Esimerkiksi osa ihmisistä saa sienistä vatsavaivoja, koska sienisokeria hajottavaa trehalaasientsyymiä ei kaikilla ihmisillä muodostu. (20, s. 55.)
Kukin entsyymi vaikuttaa yleensä vain tietyn molekyylin tai yhdisteen eli substraatin toimintaan (2). Entsyymi-substraattispesifisyydellä tarkoitetaan sitä, että jokaisen reaktion käynnistymiseen vaikuttaa oma entsyymi (13). Kaikilla entsyymeillä on myös oma tietty tehtävänsä. Entsyymejä onkin verrattu työkaluihin: tietyssä tehtävässä toimii paremmin siihen suunniteltu työkalu kuin monitoimityökalu. (15.)
Biologisten reaktioiden käynnistymiseen tarvitaan yleensä runsaasti energiaa (aktivaatioenergia), joten ne alkavat hitaasti. Entsyymit helpottavat reaktioiden käynnistymistä vähentämällä tarvittavan aktivaatioenergiaan määrää ja nopeuttavat reaktioita eli toimivat katalyytteinä. [Kuva 1] Entsyymit eivät kuitenkaan itse kulu reaktiossa. Tästä syystä entsyymeitä tarvitaan vain pieniä määriä, ja niiden voidaan sanoa olevan tehokkaita. (1; 2; 13, s. 19; 20, s. 53-54.)
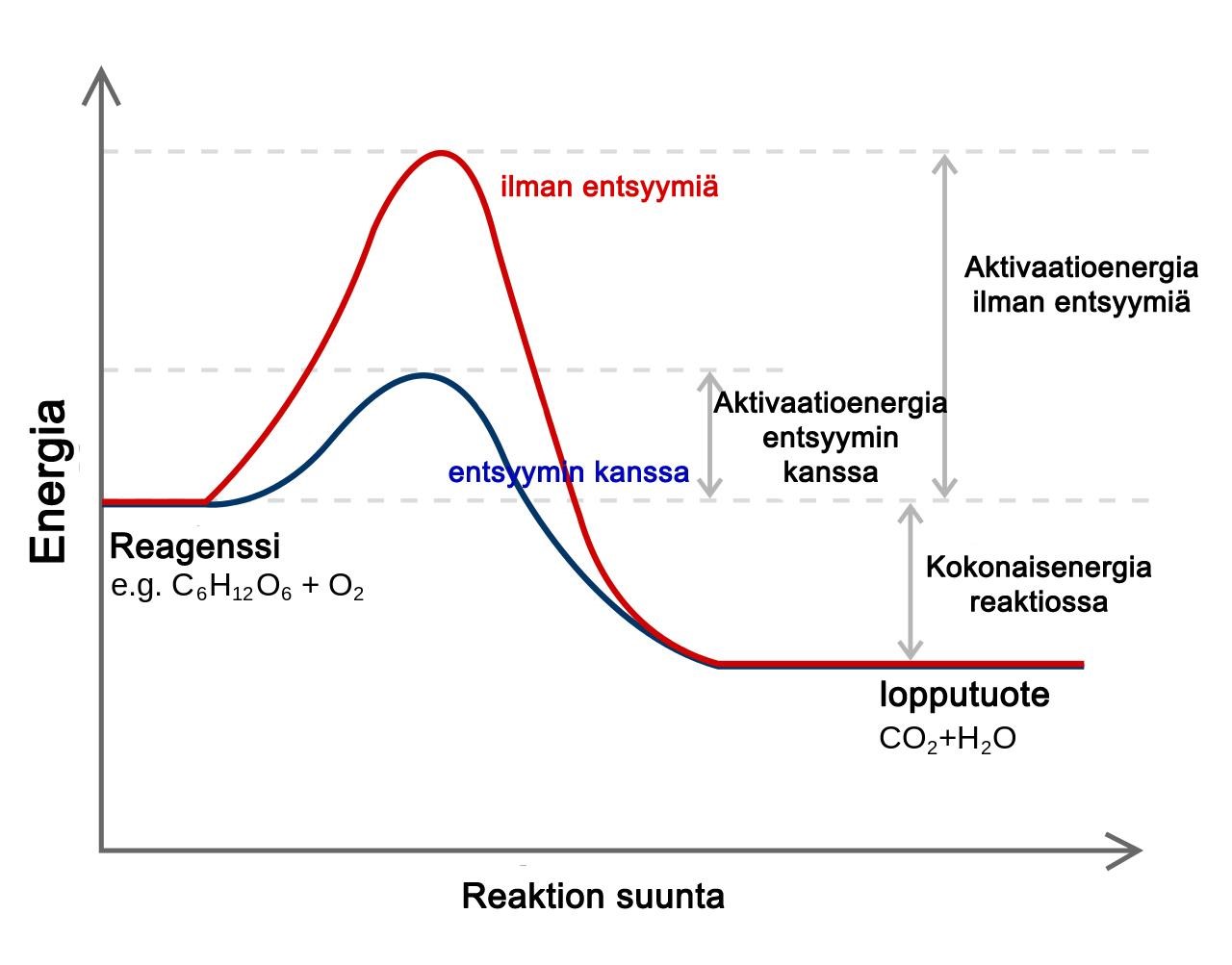
Kuva 1: Aktivaatioenergia soluhengityksessä. Tekijä: Jerry Crimson Mann 2006, muokannut: Henna Helén, Lisenssi: CC, GFDL
Entsyymien rakenne on yleensä kaksiosainen. Suurimman osan entsyymistä muodostaa proteiiniosa (eli apoentsyymi). (13, s. 20.) Osa entsyymeistä kykenee toimimaan vasta, kun niihin on kiinnittynyt erillinen osa, kofaktori (20, s. 54; 21). Kofaktorit voivat olla epäorgaanisia metalli-ioneja tai orgaanisia yhdisteitä, kuten suurikokoisia koentsyymejä, esimerkiksi vitamiineja (13, s. 20; 21; 24). Ne aktivoivat entsyymejä muuttamalla entsyymin aktiivisen alueen eli kiinnittymiskohdan substraatille sopivaksi (13, s. 20; 20, s. 54). [Kuva 2]

Kuva 2: Substraatin sitoutuessa entsyymi muuttaa muotoaan. Heksokinaasientsyymin aktiivinen alue merkitty sinisellä, substraatit (ATP ja 6-hiilinen sokeri) mustalla ja kofaktori (magnesium ioni) keltaisella. Tekijä: Thomas Shafee 2015, Lisenssi: CC
Reaktiossa substraatti kiinnittyy entsyymin aktiiviselle alueelle, syntyy entsyymi-substraatti-kompleksi, ja kemiallinen reaktio käynnistyy (3; 13, s. 20; 17, s. 29). Entsyymi ja substraatti voivat liittyä toisiinsa, koska substraatin sitoutuminen muuttaa entsyymin aktiivisen alueen muotoa (3; 22). Reaktion tuloksena substraatti muuttuu toiseksi aineeksi, lopputuotteeksi. Tällä tavoin substraatti voidaan pilkkoa tai liittää toiseen aineeseen. Reaktion loputtua entsyymi irtoaa lopputuotteesta. (3; 13, s. 20; 17, s. 29.) Entsyymit katalysoivat usein peräkkäisiä reaktioita. Tällöin edellisen reaktion lopputuote on seuraavan reaktion substraatti. (20, s. 54.)
Entsyymit toimivat esimerkiksi solujen aineenvaihdunnassa anabolisten (rakentavien) ja katabolisten (hajottavien) reaktioiden katalyytteinä. Anabolisessa reaktiossa entsyymi katalysoi substraatin rakentumista yksinkertaisemmista yhdisteistä monimutkaisemmiksi yhdisteiksi. Esimerkiksi kasvien pimeäreaktio, jossa glukoosimolekyylit rakentuvat pitkäksi ketjuksi ja muodostavat tärkkelystä. Anaboliset reaktiot sitovat energiaa. Kataboliset reaktiot ovat entsyymien hajottavia reaktioita. Niissä entsyymi katalysoi monimutkaisten yhdisteiden hajottamista yksinkertaisemmiksi yhdisteiksi. Soluhengitys on esimerkki katabolisesta reaktiosta. Siinä glukoosi hajoaa vedeksi ja hiilidioksidiksi ja samalla vapautuu energiaa solun käyttöön. (7, s. 66.) [Kuva 3]

Kuva 3: Entsyymin kiinnittyminen substraattiin ja muutos lopputuotteeksi. Anabolinen ja katabolinen reaktio. Tekijä: Henna Helén 2018, Lisenssi: public domain
Entsyymien toiminnan tehokkuuteen vaikuttavat kaikki reaktioon osallistuvat ja ulkopuoliset tekijät, mm. entsyymin ja substraatin konsentraatio, happamuus, lämpötila, sekä aktivaattorit ja inhibiittorit. (10; 11; 12 ;13, s. 21; 21; 24) Ne voivat kiihdyttää, hidastaa tai estää entsyymin toiminnan kokonaan (13, s. 21).
Useimmissa eliöissä syntyy soluhengityksen sivutuotteena vetyperoksidia. Se on haitallista soluille, joten siitä pitää päästä nopeasti eroon. (12.) Tästä syystä soluissa on katalaasientsyymiä, joka hajottaa vetyperoksidia vedeksi ja hapeksi (12; 13, s.22). Hajotettavan vetyperoksidisubstraatin määrä eli konsentraatio vaikuttaa reaktion voimakkuuteen, joka on alkuvaiheessa nopea ja tuottaa paljon happea, mutta pysähtyy vähitellen substraatin vähentyessä. (13, s. 22.)
Kukin entsyymi toimii parhaiten juuri sille optimaalisissa olosuhteissa. Entsyymin toiminta vaikeutuu tai estyy epäsopivassa lämpötilassa tai pH:ssa. Useimmat solujen entsyymit toimivat parhaiten pH-neutraalissa ympäristössä, kuten esimerkiksi syljessä tärkkelystä pilkkova amylaasientsyymi. Ruuansulatuskanavan eri osissa esiintyy entsyymejä, jotka toimivat parhaiten happamissa tai emäksisissä olosuhteissa. Mahalaukussa happamassa nesteessä (pH 2) maharauhasen erittämä pepsiini pilkkoo ruuan proteiineja peptideiksi ja aminohapoiksi. Ruokasulan edettyä pohjukaissuoleen proteiinien pilkkomista jatkaa haiman erittämä trypsiini, jonka ihanteellinen pH on 8. Haima erittää monia ruoka-aineita hajottavia entsyymejä. (10; 11; 12; 17) [Kuva 4]
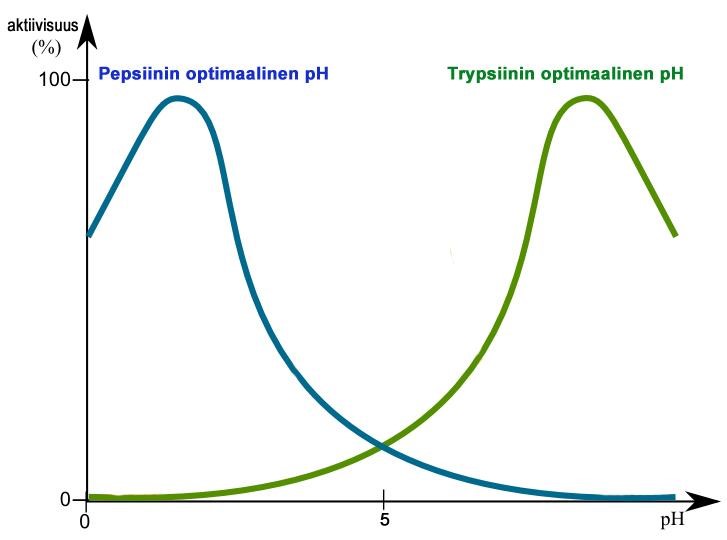
Kuva 4: Happamuuden vaikutus pepsiini- ja trypsiinientsyymien toimintaan. Tekijä: Gal M. 2007, muokannut: Henna Helén, Lisenssi: CC, GFDL
Esimerkiksi liian kuumassa ympäristössä entsyymi denaturoituu eli menettää kolmiulotteisen rakenteensa, jolloin se ei ole enää toimintakykyinen. Kuumennuksessa proteiinin kolmiulotteista rakennetta ylläpitävät rikkisidokset katkeavat ja vetysidokset järjestäytyvät uudelleen (eli rakenne muuttuu). Proteiinien denaturoitumisen korkeassa lämpötilassa voi huomata esimerkiksi ruuan valmistuksessa. Kananmunan paistaminen tai keittäminen saa munanvalkuaisen proteiinimolekyylit liittymään toisiinsa ja sitomaan vettä, jolloin valkuaisen koostumus muuttuu sakeaksi. Ihmisen entsyymit toimivat parhaiten +37 ºC:n lämpötilassa, ja yli 42ºc:n kuume johtaa denaturoitumisen vuoksi kuolemaan. Kuumissa lähteistä elävien bakteerien ja arkeonien entsyymit puolestaan toimivat parhaiten lähes +80ºC:ssa. Alhaisessa lämpötilassa entsyymien toiminta hidastuu, mutta ei lopu kokonaan edes pakkasasteiden puolella. Siksi pakastetut ruoka-aineet eivät säily ikuisesti. (7; 13, s. 22; 14.)
Tietyt kemialliset aineet aktivoivat tai ehkäisevät entsyymien toimintaa. Solut myös tuottavat itse näitä aktivaattoreita ja inhibiittoreita. Entsyymien toimintaa kiihdyttäviä aineita kutsutaan aktivaattoreiksi. Aktivaattoreita ovat esimerkiksi monet kofaktorit ja koentsyymit. (24.)
Inhibiittorit ovat aineita, jotka estävät entsyymin aktiivisuutta (7). Inhibiittorit voivat estää entsyymin toiminnan tilapäisesti tai pysyvästi (13, s. 21). Ne voivat olla joko soluille haitallisia vierasaineita tai solun omia luontaisia inhibiittoreita. Esimerkiksi entsyymireaktion lopputuote voi toimia luontaisena inhibiittorina estäen entsyymin toiminnan silloin, kun lopputuotetta on tuotettu riittävästi. Näin solun sisäiset kemialliset reaktiot pysyvät tasapainossa. (7.)
Inhibiittori voi estää entsyymin toiminnan sitoutumalla itse entsyymin aktiiviselle alueelle (kilpaileva inhibitio) tai muuttamalla entsyymimolekyylin muotoa (ei-kilpaileva inhibitio). Kilpaileva inhibiittori sitoutuu entsyymin aktiiviselle alueelle, jolloin substraatti ei voi sitoutua siihen. (7.) [Kuva 5]

Kuva 5: Kilpaileva inhibitio. Tekijä: Jerry Crimson Mann 2006, muokkaus: Henna Helén, lisenssi: public domain
Ei-kilpaileva inhibiittori sitoutuu erilliseen sitoutumiskohtaan (allosteerinen kohta) entsyymissä. Inhibiittorin sitoutuminen aiheuttaa rakenteellisen muutoksen entsyymissä, mikä estää substraatin sitoutumisen. (7) [Kuva 6] Tätä kutsutaan entsyymin allosteeriseksi säätelyksi ja se voi olla entsyymiin sitoutuneesta molekyylistä riippuen joko inhiboiva tai aktivoiva reaktio (23). Allosteeriset säätelijät ovat lupaavia entsyymien aktiivisuuteen vaikuttavia lääkeaineita, koska ne ovat hyvin spesifisiä, toisin kuin aktiiviseen keskukseen sitoutuvat inhibiittorit (16).
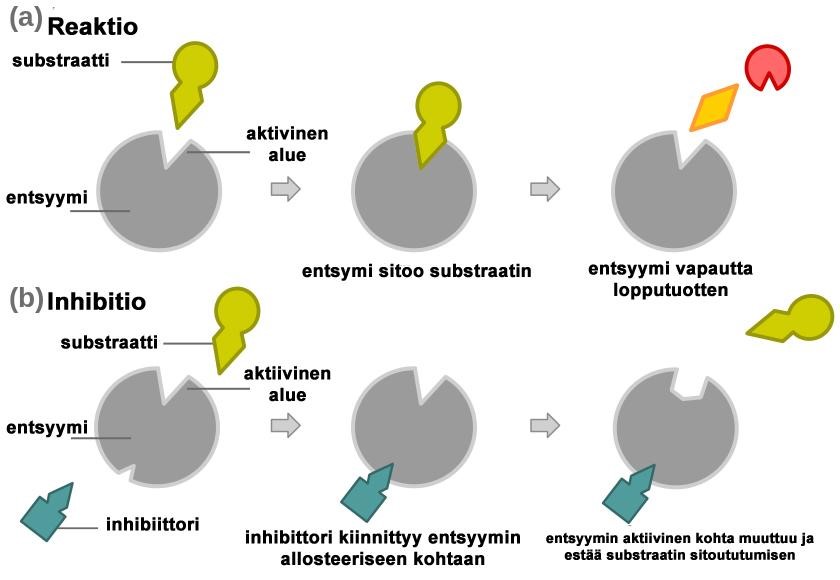
Kuva 6: Ei kilpaileva inhibitio, allosteerinen säätely. Tekijä: Jerry Crimson Mann 2009, muokannut: Henna Helén, Lisenssi: public domain
Esimerkiksi monet lääkeaineet vaikuttavat entsyymien inhibitioreaktioihin. Kivun hoito perustuu nykyisin pääasiassa tulehduskipulääkkeisiin, joiden vaikutusmekanismina on COX-entsyymien toiminnan ja prostanoidien synteesin esto. (6.) Osa antibiooteista toimii bakteerien inhibiittoreina kiinnittymällä niiden aineenvaihdunnassa toimivien entsyymien aktiiviseen kohtaan (kilpaileva inhibitio) (7). Myös monet myrkyt vaikuttavat inhiboimalla entsyymejä. Esimerkiksi syanidi sitoutuu mitokondrioiden sytokromi-c-oksidaasi-entsyymiin, denaturoi sen ja katkaisee soluhengityksen elektroninsiirtoketjun, joka johtaa solun energiantuotannon loppumiseen (8; 9).
Entsyymejä hyödynnetään monissa arkielämän tuotteissa ja niiden valmistuksessa. Esimerkiksi juuston valmistuksessa maito juoksutetaan kymosiinientsyymin avulla, ja vähälaktoosisista maitotuotteista laktoosia on pilkottu laktaasientsyymin avulla. (12.) Hammastahnaan ja purukumiin voidaan lisätä entsyymejä pahanhajuista hengitystä ehkäisemään, ja hiusväriin värin kiinnittymistä helpottamaan (19, s. 374-375). Monet pyykinpesuaineet sisältävät proteaasi-, amylaasi- ja lipaasientsyymejä, jotka hajottavat likatahrojen proteiineja, tärkkelystä ja lipidejä. Entsyymit toimivat hyvin eri lämpötiloissa, ja erilaisten entsyymien ansiosta voidaan käyttää alhaisempaa pesulämpötilaa sekä lyhyempää pesuohjelmaa. Kaikissa pesuaineissa ei kuitenkaan ole entsyymejä. Esimerkiksi villa ja silkki ovat proteiinipitoisia, ja proteaaseja sisältävä pesuaine harjoittaisi niiden kuitujen rakenteen. (12.)
Suuri osa kaupallisesti saatavilla olevista entsyymeistä on peräisin ja tuotetaan sienissä. Useimmat niistä on tarkoitettu elintarvike- ja pesuaineteollisuuden käyttöön. (19, s. 374.) Sienten kykyä hajottaa luonnonpolymeereja solunulkoisten entsyymien avulla hyödynnetään kasvibiomassojen muokkauksessa, esimerkiksi muovien raaka-aineena ja polttoaineena käytettävän etanolin valmistuksessa (19, s. 372, 379-381).
Entsyymejä hyödynnetään myös paperin valmistuksessa ja puuvillakankaiden käsittelyssä, sekä tekstiiliteollisuuden tuottamien väriaineiden puhdistuksessa jätevesistä. (19, s. 373-375, 382.) Parhaillaan tutkitaan sienten entsyymejä lääkkeiden kehitystyössä, pilaantuneen maan kunnostuksessa, sekä entsyymien vaikutusta jätevesien ympäristöä uhkaaviin yhdisteisiin ja lääkejäämiin (19, s. 374, 376, 382-383).
Aktiivinen alue, aktiivinen keskus – substraatin (ja kilpailevan inhibiittorin) sitoutumiskohta entsyymissä.
Aktivaatioenergia – reaktion käynnistymiseen vaadittavan energian määrä.
Aktivaattori – entsyymin toimintaa aktivoiva aine (engl. activate)
Allosteerinen kohta – ei-kilpailevan inhibiittorin sitoutumiskohta entsyymissä.
Anabolia – rakentava reaktio, jossa kemiallista energiaa sitoutuu orgaaniseen yhdisteeseen.
Denaturoituminen – Proteiinien rakenteen muutos, jossa toimintakyky häviää. Tämä voi tapahtua esimerkiksi liian lämpimässä, happamassa, tai suolapitoisessa ympäristössä.
Entsyymi-substraatti-kompleksi, ES-kompleksi – entsyymin ja substraatin muodostama kokonaisuus.
Entsyymi-substraattispesifisyys – Kukin entsyymi vaikuttaa tietyn substraatin toimintaan.
Inhibiittori – entsyymin toimintaa ehkäisevä aine (engl. inhibit)
Katabolia – kemiallista energiaa vapauttava, hajottava reaktio.
Katalyytti – aktivaatioenergiaa pienentävä, kemiallista reaktiota nopeuttava aine.
Kofaktori – Aine jota entsyymi tarvitsee, mutta joka ei osallistu reaktioon. Kofaktori muodostaa toimivan entsyymin yhdessä proteiiniosan kanssa.
Koentsyymi – suurikokoinen orgaaninen kofaktori, esimerkiksi vitamiini.
Lopputuote – Substraatista muodostuva, entsymaattisen reaktion päättyessä valmistuva aine.
Proteiiniosa eli apoentsyymi – Muodostaa suurimman osan entsyymistä.
Substraatti – Molekyyli tai yhdiste, jota rakennetaan tai hajotetaan entsymaattisessa reaktiossa. Kiinnittyy entsyymin aktiiviselle alueelle.
Juuston valmistus, opiskelijoiden työohje
Juuston valmistus, opettajan työohje
Tekijät: Ronja Hyppölä, Petteri Saarela & Matias Järvinen
Tämä blogipostaus sisältää opetusmateriaalia lukion BI2-kurssille Ekologia ja ympäristö. Materiaali käsittelee biodiversiteettiä, ja sisältää ”oppikirjan” kappaleen aiheesta.
Biodiversiteetti eli luonnon monimuotoisuus tarkoittaa elollisen luonnon erilaisuutta ja monipuolisuutta. Biodiversiteetillä on kolme tasoa: lajin sisäinen eli geneettinen monimuotoisuus, lajimonimuotoisuus ja ekosysteemien monimuotoisuus.
Geneettinen monimuotoisuus ilmenee jokaisella lajin yksilöllä ainutlaatuisena perimänä. Jokainen yksilö poikkeaa muista saman lajin yksilöistä geneettiseltä koostumukseltaan. Vaihtelua esiintyy esimerkiksi kasvien kylmänsietokyvyssä, kasvunopeudessa tai eläinten käyttäytymisessä. Geneettinen monimuotoisuus on tärkeää lajin sopeutumisen ja säilymisen kannalta. Yksilöllinen muuntelu mahdollistaa lajin sopeutumisen, kun laji leviää uusille alueille tai jos elinympäristö muuttuu. Monimuotoisuus suojaa populaatioita myös taudeilta, tuholaisilta ja saasteilta.

Kuva: Wikimedia Commons.
Lajimonimuotoisuus tarkoittaa lajien runsautta ekosysteemissä tai jollakin tietyllä alueella. Sademetsissä ja koralliriutoilla lajimonimuotoisuus on runsainta. Alueita, joilla monimuotoisuus on erityisen suurta, kutsutaan monimuotoisuus- eli hotspot-keskuksiksi. Myös vierekkäisten ekosysteemien raja-alueilla on runsaasti lajeja niin sanotulla reunavaikutusalueella. Suomessa lajimonimuotoisuus on suurinta lehdoissa ja lettosoilla. Jokaisella lajilla on oma paikkansa ekosysteemissä. Esimerkiksi monet hyönteiset pölyttävät kasveja ja toiset eläimet voivat auttaa kasveja levittäytymään syömällä ja ulostamalla niiden siemeniä. Jos ekosysteemissä lajimäärä muuttuu lajin katoamisen tai uuden lajin leviämisen takia, vaikutukset voivat ulottua koko ekosysteemiin.
Ekosysteemien monimuotoisuus on laajin monimuotoisuuden taso. Sillä tarkoitetaan erilaisten elinympäristöjen eli biotooppien monimuotoisuutta. Biotooppien monimuotoisuutta voidaan tutkia jollain tietyllä alueella tai koko maapallolla. Ekosysteemien monimuotoisuuteen vaikuttavat muun muassa lämpö, valo, maaperän ravinnepitoisuus ja kosteusolosuhteet. Maapallolla on 10 eri suurekosysteemiä, jotka ovat ekosysteemien muodostamia kokonaisuuksia. Maapallon 10 suurekosysteemiä ovat sademetsät, savannit, aavikot ja puoliaavikot, nahkealehtiset kasvit, subtrooppiset sademetsät, arot, lehtimetsät, havumetsät, tundrat ja vuoristot. Ekosysteemien rajat eivät ole tarkkoja, ja ekosysteemit vaikuttavat toisiinsa.

Kuva: Wikimedia Commons. Vertaile sademetsän ja tundran biotooppeja.
Luonnon monimuotoisuus on jakautunut maapallolla epätasaisesti, sillä abioottiset eli elottomat tekijät, kuten valo, lämpö ja sademäärä vaihtelevat alueittain. Kunkin alueen abioottiset tekijät määrittävät, millainen ekosysteemi sinne muodostuu. Maapallo on jaettu suurekosysteemeihin eli biomeihin, joita ovat esimerkiksi pohjoiset havumetsät, aavikot ja trooppiset sademetsät.
Alueiden biodiversiteettiin vaikuttavia tärkeimpiä abioottisia tekijöitä ovat alueen pinta-ala, lämpötila, auringon säteily ja sadanta. Pinta-alaltaan laajalla alueella on paljon tilaa erilaisille eliölajeille ja -yhteisöille. Kun luonnonolot pysyvät vakaina, esimerkiksi vuoden ympäri lämpimissä ja kosteissa sademetsissä, eliöt eivät altistu muutoksista aiheutuvalle stressille. Pidemmällä aikavälillä vakaiden alueiden biodiversiteettiä vahvistaa se, että lajit ovat ehtineet aikojen saatossa sopeutumaan tiettyihin pysyviin paikkoihin eliöyhteisössä eli ekolokeroihin. Vakiintuneet olosuhteet ja eristyminen muista alueista voi johtaa myös ainutlaatuisiin endeemisiin lajistoihin, kuten Galapagossaarilla.
Suurin osa maapallon ekosysteemeistä perustuu auringon säteilyn energiaan, jota kasvit ja levät hyödyntävät yhteyttäessään. Maapallolla eniten auringon säteilyä saadaan päiväntasaajalla, vähiten napa-alueilla. Tästä johtuen perustuotanto ja ekosysteemien lajimäärät vähenevät napoja kohti edetessä. Maailman lajirikkaimpia alueita ovat trooppiset sademetsät ja koralliriutat.
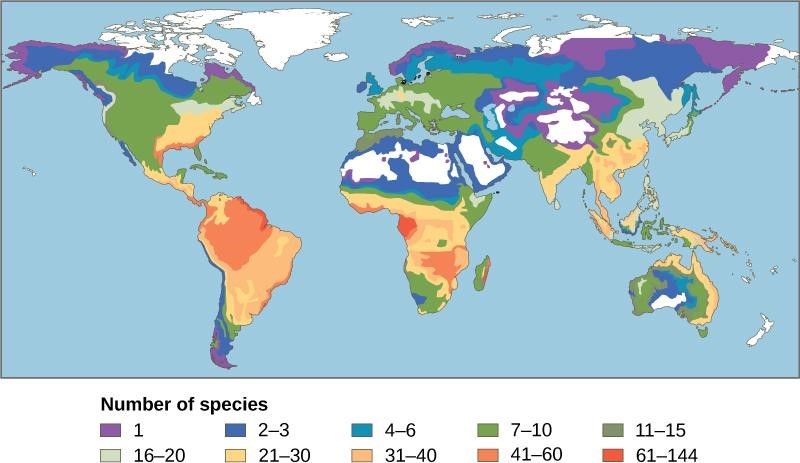
Kuva: Wikimedia Commons. Kuva sammakkoeläinten lajimääristä.
Biologian sanakirjassa biodiversiteetti on määritetty seuraavasti: “elollisen luonnon monimuotoisuus; tarkoittaa eliölajien perinnöllistä muuntelua, eliöyhteisön lajiston monimuotoisuutta ja erilaisten eliöyhteisöjen kirjoa.” Miten monimuotoisuutta voidaan mitata tai minkälaisia keinoja biodiversiteetin arvioimiseksi käytetään?

Kuvat: Wikimedia Commons. Eliökunnan diversiteettiä.
Yleisin mittaamistapa on jonkin paikan tai alueen laji-lukumäärän laskeminen. Lähtökohtaisesti tarkka laskeminen on kuitenkin hankalaa, sillä biodiversiteetin tasoja on useita, lajin sisäisistä erilaisiin eliöryhmiin. Sen lisäksi laskemisen toteuttaminen kentällä on haasteellista; jotkin lajit ovat liian pieniä tullakseen nähdyiksi, toiset taas eivät välttämättä ole laskentahetkellä paikalla vaikka normaalisti habitaattia asuttavatkin. Kaikkiaan biodiversiteetin mittaaminen on lähes aina vain suuntaa antavaa oikeassa luonnossa. Mittaamista ja biodiversiteettien vertailua voidaan kuitenkin tehdä, kun ensin päätetään selkeästi mitattavan biodiversiteetin taso, mitä eliöryhmiä huomioidaan, millä perusteilla eri lajeja painotetaan jos se on habitaatin kannalta oleellista ja millä tarkkuudella laskenta pyritään tekemään.
Joskus huomio kiinnitetään lajien tärkeyteen. Jotkin lajit ovat ns. avainlajeja, joiden läsnäolo vaikuttaa muihin lajeihin. Avainlajit mahdollistavat muiden lajien elämän kyseisessä habitaatissa. Usein tällaisia avainlajeja painotetaan laskennoissa niiden tärkeyden takia. Joitakin vain vähäisiä elinympäristön muutoksia sietäviä lajeja kutsutaan indikaattori- eli ilmentäjälajeiksi. Indikaattorilajien tai -ryhmien läsnäolo kertoo, että habitaattiin vaikuttaa jokin biologinen tai fysikaalis-kemiallinen tekijä, esimerkiksi pohjavesi, joka ei kuitenkaan välttämättä näy maisemassa päällepäin oikeana vesistönä. Jos tämä tekijä puuttuu, indikaattorilaji ei enää viihdy paikalla. Jotkin indikaattorilajit voivat ilmentää että habitaatissa esiintyy myös jotain toista tiettyä lajia; lajit ovat siis yhteydessä toisiinsa.

Kuva: Wikimedia Commons. Indikaattorilaji, rahkasammalet kertovat veden happamuudesta.
Monimuotoisuutta voidaan mitata myös vertailemalla lajien ainutlaatuisuutta tai paikallisten kotoperäisten lajien määrää. Hyvin äärimmäisissä elinympäristöissä asuvat lajit ovat ainutlaatuisempia kuin lajit joita tavataan monenlaisissa ympäristöissä. Toisaalta jos laji on sukunsa ainoa edustaja, on se ainutlaatuisempi, kuin laji jolla on useita läheisiä sukulaislajeja. Kotoperäiset eli endeemiset lajit asuttavat vain tiettyä aluetta ja ovat sopeutuneet elämään juuri siellä; ne ovat sieltä kotoisin. Kotoperäisten lajien esiintyminen painottaa kyseisen alueen tai habitaatin tärkeyttä biodiversiteetin säilymisen kannalta.
Monimuotoisuutta voidaan arvioida myös vertailemalla kahta eri aluetta tai habitaattia ja niiden lajien välistä erilaisuutta. Esimerkiksi järvi, jossa asuu ahvenia ei ole yhtä monimuotoinen kuin järvi, jossa on ahventen lisäksi haukia. Kuitenkin jos pelkkien ahvenien asuttaman järven ahvenet ovat geneettisesti monimuotoisia, lisää se myös koko järven monimuotoisuutta. Järvi voi myös luoda sen alueelle erilaisia habitaatteja, esimerkiksi erilaisia rantoja kallioisista hiekkaisiin, tai reheviä- ja karuja saaria. Mitä enemmän näitä erilaisia elinympäristöjä on, sen monimuotoisempi järven alue on luultavasti myös lajistoltaan.
Yleisesti diversiteetistä puhuttaessa käytetään termejä alfa-, beeta- ja gammadiversiteetti, mitkä viittaavat diversiteettien maantieteelliseen tarkasteluun.
Alfadiversiteetti kuvaa alueellista monimuotoisuutta, beetadiversiteetti taas alueiden välistä diversiteettiä. Gammadiversiteetti on maantieteellisesti laajan alueen monimuotoisuus; usean alfadiversiteetin yhteenlaskettu monimuotoisuus.
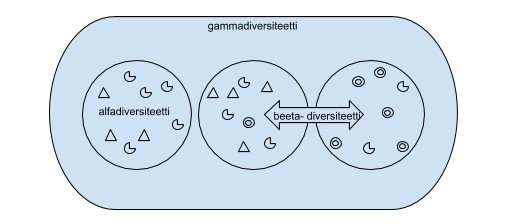
Kuva: Ronja Hyppölä. Maantieteellinen alfa-, beta-, ja gammadiversiteetti.